Molecular & Biochemical Parasitology 131 (2003) 77–81
The H region HTBF gene mediates terbinafine resistance
Julio F.M. Marchini , Angela K. Cruz , Stephen M. Beverley , Luiz R.O. Tosi
a Departamento de Biologia Celular e Molecular e Bioagentes Patogˆenicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo,Ave. Bandeirantes, 3900, 14049-900 Ribeirão Preto—SP, Brazil
b Department of Molecular Microbiology, Washington University Medical School, 660 South Euclid Ave., St. Louis, MO 63110, USA
Received 15 March 2003; received in revised form 23 June 2003; accepted 30 June 2003
Keywords: Leishmania; Terbinafine resistance; H region; Gene amplification; In vitro transposition
Leishmania spp. are the causative agent of leishmania-
sistant to heavy metals (arsenite and antimonials) through
sis, a disease that has an estimated global prevalence of 12
a still unclear mechanism The alternative pteridine
million cases The disease has a broad spectrum of clin-
reductase PTR1 confers resistance to methotrexate, and has
ical presentations, from skin and subcutaneous lesions to
been shown to limit pathogenesis within the mammalian
visceral disease. The mainstream treatment for leishmania-
host L. major subject to drug pressure may become
sis consists of parenteral administration of pentavalent anti-
resistant to terbinafine through H region amplification, as
monials, which may result in dose-cumulative side effects.
observed in cell lines SF-R30 (selected in terbinafine) and
Amphotericin B is an even more toxic and less effective al-
PQ-R10 (selected in primaquine) erbinafine is an anti-
ternative. For the treatment of complex cases, inhibitors of
fungal drug that is capable of inhibiting squalene-epoxidase
the sterol biosynthesis pathway, such as terbinafine, offer an
a key enzyme in the biosynthesis of ergosterol, an
attractive possibility as they target parasite-specific physio-
essential component of the cell membrane. Terbinafine tox-
icity thus arises from ergosterol deficiency and squalene
The study of the ability of this organism to evade
accumulation the genes mediating resistance
chemotherapy is important not only to understand different
to other agents that induce H region amplification have
aspects of this protozoan’s biology but also to help the
been identified, terbinafine resistance has not been mapped
design of effective treatments. Gene amplification is a com-
mon mechanism used for survival in Leishmania strains
The effects of terbinafine on the parasite growth are com-
selected for resistance to drugs such as methotrexate
plex and were shown to elicit resistance. The identification
Leishmania major selected in methotrexate, primaquine,
of a terbinafine-resistance gene present within the H region
or pentavalent antimonials derive resistance to these drugs
took advantage of a strategy based upon gene inactivation
through the amplification of a 48 kb locus in chromosome
through insertional mutagenesis and its correlation with loss
23, the H region. P-glycoprotein A (PGPA) and Pteridine
of phenotype. Considering the 48 kb span of the H locus, the
reductase 1 (PTR1) are the two genes related to drug resis-
resistance phenotype was constrained to a smaller fragment
tance that have been identified within the H region. PGPA
in order to reduce the number of genes to be inactivated by
is an ATP-binding cassette which renders the parasite re-
the insertional mutagenesis protocol. The H region had al-ready been divided into four fragments cloned into the shut-tle vector pSNAR Terbinafine resistance was assessed
Abbreviations: PCR, polymease chain reaction; GFP, green fluores-
cent protein; RTPCR, reverse transcriptase polymerase chain reaction;
using not only L. major transfectants bearing each of the
PFGE, pulse field gel electrophoresis; YIP, YPT interacting protein; PGPA,
four clones, but also the SF-R30 strain and the wild-type cell
P-glycoprotein A; PTR1, Pteridine reductase 1; ORF, open reading frame;
line LT252 as positive and negative controls, respectively.
MOPS, 3-[N-morpholino] propane-sulfonic acid; HTBF, H region asso-
The fold resistance at EC50 for these lines is presented in
ciated terbinafine resistance; SNARE, soluble N-ethylmaleimide-sensitive
Transfectants bearing the H region subfragments
∗ Corresponding author. Tel.: +55-16-602-3117; fax: +55-16-633-1786.
H1, H2, and H3 were not different from the wild-type cell
E-mail address: [email protected] (L.R.O. Tosi).
line. However, the fold resistance exhibited by transfectant
0166-6851/$ – see front matter 2003 Elsevier B.V. All rights reserved. doi:10.1016/S0166-6851(03)00174-9
J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 1. Delimitation of the terbinafine-resistance gene within the H region. (A) SacI fragments of H4 (T1, T2, and T4) were subcloned into pELHYGand tested for terbinafine resistance; plus and minus signs represent resistance and sensitivity, respectively; black arrows represent inverted repeats (T3aand T3b); S = SacI site; sequencing of T1 revealed two complete open reading frames indicated by white arrows; the arrows on HTBF represent theplacement of mariner transposon insertion events; the insertion named 82 was used to originate the L82 transfectant. (B and C) Fold resistance of L. major cell lines and transfectants at EC50 for terbinafine relative to that of the wild-type LT252, CC1 clone (MHOM/IR/84/LT 252); log-phase cellswere seeded at a cell density of 105 cells ml−1 into drug-free medium as well as in media containing terbinafine ranging from 2 to 8 g ml−1; thedrug-free culture growth was monitored daily until it reached late-log phase (0.8 to 1.0 × 107 cells ml−1), when the terbinafine EC50 was determinedas the concentration that decreased the cell growth rate by 50%; mean and S.D. are shown; ∗Values significantly higher than the wild-type value byStudent’s t test (P < 0.05); ∗∗Values significantly higher than all other lines and transfectants by Student’s t test (P < 0.05). Terbinafine resistantcell lines were SF-R30 and PQ-R10 stable transfectants were initially selected on 32 g ml−1 of hygromycin B and increased to 500 g ml−1 inindicated transfectants by doubling steps; transfectants were named according to Clayton et al The fold-resistance is the ratio of terbinafine EC50sfor experimental and LT252 cells, measured in the same experimenta.
H4 (LT[pSNAR8H4]) was comparable to that of the resis-
ent mechanisms of terbinafine resistance in addition to the
tant cell line PQ-R10 and significantly higher than that ob-
served for wild-type cells (P < 0.05). Although fold re-
In order to limit the locus that has imparted terbinafine re-
sistance is typically low (about two-fold), it is consistently
sistance to Leishmania, the H4 fragment was restricted with
found in terbinafine resistant lines regardless of the selec-
SacI, which generated five fragments (Two of the
tive force leading to H region amplification. The cell lines
resulting fragments encompass the inverted repeats present
selected in terbinafine are an exception in which the fold
in the right end of the H locus. The remaining fragments
resistance is significantly higher Different mechanisms
were subcloned into shuttle vectors pELHYG or pELHYGII
might play a role in bringing about terbinafine resistance. In
and transfected into the parasite. L. major transfectants T1
fact, Leishmania resistance to sterol biosynthesis inhibitors
(LT[pELHYG32T1]) and T4 (LT[pELHYGII32T4]) were iso-
can be achieved by the expression of many loci in-
lated and kept under 32 g ml−1 hygromycin B. The concen-
cluding the H locus, as shown in SF-R30 and PQ-R10 lines.
tration of hygromycin B in culture was raised to 500 g ml−1
Therefore, we cannot exclude the possibility that terbinafine
in doubling steps so as to increase the plasmid copy num-
selected cell lines, such as SF-R30, have developed differ-
ber. The transfectants T1500 (LT[pELHYG500T1]) and T4500
J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
(LT[pELHYGII500T4]) allowed the analysis of the effect of
prenylated to a geranylgeranyl group that mediates attach-
this phenomenon on the terbinafine-resistance phenotype.
ment to membranes pool of YPT exists in the cytosol,
Control cell lines, as well as the different transfectants noted
chaperoned by GDP dissociation inhibitor (GDI). Interac-
above, were used in a terbinafine resistance-assay (
tion with YIP1 promotes the dissociation of the YPT/GDI
The fold resistance of transfectants T4500 and T132 was not
heterodimer, allowing YPT to reinsert into membranes
different from that observed for the wild-type cells. Nev-
A YPT homologue gene has already been identified in L. ma-
ertheless, the transfectant T1500 exhibited a fold resistance
jor and its product has been localized in the parasite Golgi
that was significantly higher than that observed for wild-type
cells and comparable to that exhibited by the transfectant
The predicted HTBF is a 21 kDa protein containing four
H4. These results not only consistently linked the resistance
membrane-spanning domains, with both C and N terminus
phenotype to the H locus, but also narrowed the search for
oriented toward the cytoplasmic face of the membrane.
the resistance gene to only 2.8 kb contained in T1 fragment.
Kyte-Doolittle hydrophobicity plot analysis also indicated
Primer-island sequencing of the segment cloned into pEL-
that HTBF is potentially an integral membrane protein.
HYG revealed two complete open reading frames (ORFs)
Southern blot analysis of pulse field gel electrophoresis
within the T1 locus (accession number AY227807). ORF2
(PFGE)-separated chromosomes, as well as SacI digested
and ORF3 had typical upstream and downstream putative
genomic DNA, suggested that HTBF is present in one copy
AG trans-splicing acceptor sites and pyrimidine-rich tracks
in the chromosome 23 of L. major (data not shown).
An incomplete open reading frame, ORF1, lacking a
The transcription of HTBF was studied using Northern
5 -end was identified upstream of ORF2. Comparison with
blot analysis of total RNA extracted from Leishmania cell
the public sequence databases revealed the homology of
lines LT252 and SF-R30, as well as from transfectants
the 570 bp ORF2 to genes annotated in the human, worm,
T4500, T132, T1500, and L82. The 570 bp HTBF probe rec-
and yeast genomes. No significant sequence homology was
ognized predominantly a 1.9 kb transcript in T132, T1500,
found for 333 bp ORF1 and 360 bp ORF3.
and SF-R30, which was absent in LT252 line or transfec-
Since transfection of segment T1 clearly rendered L. ma-
tant T4500 (Although the basal level of the tran-
jor resistant to terbinafine, this fragment was selected as a
script was not detected, its expression is evident in the H
target for an insertional mutagenesis protocol. Inactivation
region-amplified cell line and in the transfectants carrying
of genes within T1 was carried out in vitro using the mariner
the intact recombinant HTBF. Increased drug pressure led
transposition system Specific pairs of primers for each
to higher transcript levels in T1500 when compared to T132.
gene within T1 were used in a colony polymease chain re-
A 5 kb transcript, which could correspond to read-through
action (PCR) protocol in order to identify useful insertion
transcription across the circular episome was also de-
events. Primers LT007 and LT008 were selected to screen
tected in transfectants carrying pELHYG-T1, but not in the
insertion events into ORF2, revealing 5 out of 96 events that
SF-R30 line. As a consequence of the insertion into HTBF,
had the transposable element inserted within the 570 bp ORF.
the transcripts detected in transfectant L82 were 2 kb larger
One of the insertions disrupting ORF2, named 82 (
than those found in T1500. HTBF transcripts were also
was transfected into L. major. The transfectant obtained, L82
detected in T1500 and SF-R30 through a RT-PCR method
(LT[pELHYG500T1::/GFP∗K82]), was used in a terbinafine
(Being a more sensitive method, the RT-PCR
resistance-assay where T1500, H4, and LT252 were the pos-
also detected the basal level of transcript in the LT252 line
itive and negative controls, respectively. The transfectant
and transfectant T4500. As expected, a 2 kb larger band
L82, in which the insertion occurred 141 bp downstream of
was also amplified from L82 total RNA. Reactions that did
ORF2 ATG, presented a fold resistance that was not differ-
not include the reverse transcriptase confirmed the absence
ent from the wild-type, indicating the loss of the terbinafine
of contaminating DNA in the template samples (data not
resistance-phenotype Therefore, the integration
of the mariner element/GFP∗K at one fourth of the putative
HTBF could mediate resistance to terbinafine through
gene length was enough to inactivate the terbinafine resis-
a general mechanism involving the increased formation
tance associated to the 2.8 kb fragment T1. ORF2 was thus
and/or the redirection of vesicles. Studies on the mecha-
named H region associated terbinafine resistance (HTBF).
nism of metal resistance in Leishmania mediated by the
The BLAST analysis of HTBF revealed that its predicted
ABC transporter PGPA suggested that this protein acts on
amino acid sequence had significant homology to the YIP1
metal-thiol conjugates PGPA is associated to intra-
protein of Saccharomyces cerevisiae. The predicted HTBF
cellular membranes in structures proposed to be part of
and the yeast protein shared 23.9% identity at the amino acid
exocytic and endocytic pathways in Leishmania
level and there was 38.4% identity between this HTBF and
findings indicated that PGPA confers resistance to arsenite
the Caenorhabditis elegans homologue. Overall, the Leish-
and antimonials by sequestration of metals into vesicles
mania HTBF shared 27.0% identity with the domain consen-
that could be exocytosed through the flagellar pocket. In
sus of YIP1 protein from different organisms (COG5080).
this scenario, HTBF would improve the process of vesicle
In the yeast, YIP1 interacts with YPT, a protein related to
trafficking and/or docking. It is noteworthy that L. tarento-
vesicle docking and trafficking YPT is a Rab/GTPase
lae H region-derived amplicons containing PGPA are likely
J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 2. Analyses of HTBF transcripts. (A) Northern analysis of total RNA in the cell lines LT252 (lane 1); SF-R30 (lane 6) and transfectantsT4500 (lane 2); T132 (lane 3); T1500 (lane 4) and L82 (lane 5); the black arrowhead indicates the ∼3.9 kb transcript in the transfectant bearingthe transposon insertion into HTBF. Total RNA from promastigote forms was extracted using TRIzol® reagent (Gibco BRL) and manipulated asdescribed elsewhere The HTBF probe was amplified by PCR with primers LT007 (5 -GCGCCCGGGCATATGCTCAACGAGGTGC) and LT008(5 -CGCGGATCCTAAATACCAACCAGA). (B) RT-PCR amplification products using HTBF primers (LT007/LT008) and total RNA from LT252 (lane1); T4500 (lane 2); T1500 (lane 4); L82 (lane 5); and SF-R30 (lane 6); the white arrowhead indicates the amplification product of 2.5 kb which representsthe insertion of element/GFP∗K into HTBF; reactions that did not include the reverse transcriptase confirmed the absence of contaminating DNA inthe template samples (data not shown); reverse transcriptase-PCR was performed with Ready-To-GoTM RT-PCR Beads (Amersham Pharmacia Biotech)according to the manufacturer’s specifications.
to carry the HTBF gene which is located only 2.2 kb
[2] Rangel H, Dagger F, Hernandez A, Liendo A, Urbina JA. Naturally
upstream of PGPA in L. major.
azole-resistant Leishmania braziliensis promastigotes are renderedsusceptible in the presence of terbinafine: comparative study with
The terbinafine interference on the ergosterol biosyn-
azole-susceptible Leishmania mexicana promastigotes. Antimicrob
thesis may result in the use of exceptional sterols and
cause membrane malfunctioning. It has been suggested that
[3] Ellenberger TE, Beverley SM. Multiple drug resistance and conser-
Ca2+-dependent exocytosis may be essential in the main-
vative amplification of the H region in Leishmania major. J Biol
tenance of cell membrane integrity as it lowers surface
[4] Callahan HL, Roberts WL, Rainey PM, Beverley SM. The PGPA
tension In fact, proteolytic toxins, which specifi-
gene of Leishmania major mediates antimony (SbIII) resistance by
cally cleave soluble N-ethylmaleimide-sensitive factor at-
decreasing influx and not by increasing efflux. Mol Biochem Parasitol
tachment protein receptors (SNARE), also block exocytosis
and inhibit membrane resealing. A calcium-dependant er-
[5] Legare D, Papadopoulou B, Roy G, Mukhopadhyay R, Haimeur A,
gosterol mutant of S. cerevisiae had already been described
Dey S, et al. Efflux systems and increased trypanothione levels in
studies of a temperature-sensitive yeast mutant
arsenite-resistant Leishmania. Exp Parasitol 1997;87:275–82.
[6] Ouellette M, Fase-Fowler F, Borst P. The amplified H circle
of YPT revealed that rescue from growth arrest could be
of methotrexate-resistant Leishmania tarentolae contains a novel
achieved by increasing extracellular Ca2+ These ob-
P-glycoprotein gene. EMBO J 1990;9:1027–33.
servations clearly indicate that increased external levels of
[7] Cunningham ML, Titus RG, Turco SJ, Beverley SM. Regulation
calcium ions can rescue either mutants of ergosterol biosyn-
of differentiation to the infective stage of the protozoan parasite
thesis or vesicle-docking pathways by triggering the mem-
Leishmania major by tetrahydrobiopterin. Science 2001;292:285–7.
[8] Callahan HL, Beverley SM. A member of the aldoketo reductase
brane repairing machinery. Thus, terbinafine interference in
family confers methotrexate resistance in Leishmania. J Biol Chem
Leishmania ergosterol biosynthesis would be supposedly
counteracted by the activation of the membrane repairing
[9] Papadopoulou B, Roy G, Ouellette M. A novel antifolate resis-
machinery through the enhanced expression of HTBF.
tance gene on the amplified H circle of Leishmania. EMBO J1992;11:3601–8.
[10] Petranyi G, Ryder NS, Stutz A. Allylamine derivatives: new class
of synthetic antifungal agents inhibiting fungal squalene epoxidase. Acknowledgements
[11] Ryder NS. Terbinafine: mode of action and properties of the squalene
This work was supported by Fundação de Amparo a
epoxidase inhibition. Br J Dermatol 1992;126(Suppl 39):2–7.
Pesquisa do Estado de São Paulo, FAPESP, 98/09805–0;
[12] Leyden J. Pharmacokinetics and pharmacology of terbinafine and
itraconazole. J Am Acad Dermatol 1998;38:S42–7.
and UNDP/WORLD BANK/WHO Special Programme for
[13] Callahan HL, Beverley SM. Heavy metal resistance: a new role for
Research and Training in Tropical Diseases; JFMM was
P-glycoproteins in Leishmania. J Biol Chem 1991;266:18427–30.
[14] Cotrim PC, Garrity LK, Beverley SM. Isolation of genes me-
diating resistance to inhibitors of nucleoside and ergosterolmetabolism in Leishmania by overexpression/selection. J Biol Chem1999;274:37723–30. References
[15] LeBowitz JH, Smith HQ, Rusche L, Beverley SM. Coupling of
poly(A) site selection and trans-splicing in Leishmania. Genes Dev
[1] WHO. Weekly epidemiological record. 2002;44:365–72. J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
[16] Tosi LRO, Beverley SM. Cis and trans factors affecting Mos1 mariner
PGPA is an intracellular metal-thiol transporter ATPase. J Biol Chem
evolution and transposition in vitro, and its potential for functional
genomics. Nucleic Acids Res 2000;28:784–90.
[23] Grondin K, Roy G, Ouellette M. Formation of extrachromosomal cir-
[17] Yang X, Matern HT, Gallwitz D. Specific binding to a novel and
cular amplicons with direct or inverted duplications in drug-resistant
essential Golgi membrane protein (Yip1p) functionally links the
Leishmania tarentolae. Mol Cell Biol 1996;16:3587–95.
transport GTPases Ypt1p and Ypt31p. EMBO J 1998;17:4954–63.
[24] Togo T, Alderton JM, Steinhardt RA. Long-term potentiation of
[18] Schimmoller F, Simon I, Pfeffer SR. Rab GTPases, directors of
exocytosis and cell membrane repair in fibroblasts. Mol Biol Cell
vesicle docking. J Biol Chem 1998;273:22161–4.
[19] Calero M, Whittaker GR, Collins RN. Yop1p, the yeast homolog of
[25] Crowley JH, Tove S, Parks LW. A calcium-dependent ergosterol
the polyposis locus protein 1, interacts with Yip1p and negatively
mutant of Saccharomyces cerevisiae. Curr Genet 1998;34:93–9.
regulates cell growth. J Biol Chem 2001;276:12100–12.
[26] Schmitt HD, Puzicha M, Gallwitz D. Study of a temperature-sensitive
[20] Cappai R, Osborn AH, Gleeson PA, Handman E. Cloning and char-
mutant of the ras-related YPT1 gene product in yeast suggests a
acterization of a Golgi-associated GTP-binding protein homologue
role in the regulation of intracellular calcium. Cell 1988;53:635–
from Leishmania major. Mol Biochem Parasitol 1993;62:73–82.
[21] Grondin K, Haimeur A, Mukhopadhyay R, Rosen BP, Ouellette
[27] Clayton C, Adams M, Almeida R, Baltz T, Barrett M, Bastien P,
M. Co-amplification of the gamma-glutamylcysteine synthetase gene
et al. Genetic nomenclature for Trypanosoma and Leishmania. Mol
gsh1 and of the ABC transporter gene pgpA in arsenite-resistant
Leishmania tarentolae. EMBO J 1997;16:3057–65.
[28] Sambrook J, Fritsch E, Maniatis T. Molecular cloning: a laboratory
[22] Legare D, Richard D, Mukhopadhyay R, Stierhof YD, Rosen BP,
manual, 2nd ed. Cold Spring Harbor: Cold Spring Harbor Laboratory
Haimeur A, et al. The Leishmania ATP-binding cassette protein
Informed Consent Laparoscopic Possible Open LapBand G. Derek Weiss, M.D., F.A.C.S. John S. Oldham, Jr., M.D. You have decided to undergo laparoscopic, possible open, LapBand placement. During the last several weeks/months, as we have prepared you for your surgery, we have provided you with complete and detailed information about the operation, as well as the other options and procedur
DEPARTMENT OF EARTH SCIENCES CONFIDENTIAL UNI VERSI TY OF C AM BRI DGE SAFETY/MEDICAL FORM FOR ALL NEW MEMBERS OF THE DEPARTMENT (PLEASE PRINT) POSITION IN DEPT …………………………………. SUPERVISOR/MANAGER ………….……….…………….…. EMAIL ………………………………………………………….ROOM NO ……………………………….�


 Molecular & Biochemical Parasitology 131 (2003) 77–81
The H region HTBF gene mediates terbinafine resistance
Julio F.M. Marchini , Angela K. Cruz , Stephen M. Beverley , Luiz R.O. Tosi
a Departamento de Biologia Celular e Molecular e Bioagentes Patogˆenicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo,
Ave. Bandeirantes, 3900, 14049-900 Ribeirão Preto—SP, Brazil
b Department of Molecular Microbiology, Washington University Medical School, 660 South Euclid Ave., St. Louis, MO 63110, USA
Received 15 March 2003; received in revised form 23 June 2003; accepted 30 June 2003
Keywords: Leishmania; Terbinafine resistance; H region; Gene amplification; In vitro transposition
Leishmania spp. are the causative agent of leishmania-
sistant to heavy metals (arsenite and antimonials) through
sis, a disease that has an estimated global prevalence of 12
a still unclear mechanism The alternative pteridine
million cases The disease has a broad spectrum of clin-
reductase PTR1 confers resistance to methotrexate, and has
ical presentations, from skin and subcutaneous lesions to
been shown to limit pathogenesis within the mammalian
visceral disease. The mainstream treatment for leishmania-
host L. major subject to drug pressure may become
sis consists of parenteral administration of pentavalent anti-
resistant to terbinafine through H region amplification, as
monials, which may result in dose-cumulative side effects.
Molecular & Biochemical Parasitology 131 (2003) 77–81
The H region HTBF gene mediates terbinafine resistance
Julio F.M. Marchini , Angela K. Cruz , Stephen M. Beverley , Luiz R.O. Tosi
a Departamento de Biologia Celular e Molecular e Bioagentes Patogˆenicos, Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo,
Ave. Bandeirantes, 3900, 14049-900 Ribeirão Preto—SP, Brazil
b Department of Molecular Microbiology, Washington University Medical School, 660 South Euclid Ave., St. Louis, MO 63110, USA
Received 15 March 2003; received in revised form 23 June 2003; accepted 30 June 2003
Keywords: Leishmania; Terbinafine resistance; H region; Gene amplification; In vitro transposition
Leishmania spp. are the causative agent of leishmania-
sistant to heavy metals (arsenite and antimonials) through
sis, a disease that has an estimated global prevalence of 12
a still unclear mechanism The alternative pteridine
million cases The disease has a broad spectrum of clin-
reductase PTR1 confers resistance to methotrexate, and has
ical presentations, from skin and subcutaneous lesions to
been shown to limit pathogenesis within the mammalian
visceral disease. The mainstream treatment for leishmania-
host L. major subject to drug pressure may become
sis consists of parenteral administration of pentavalent anti-
resistant to terbinafine through H region amplification, as
monials, which may result in dose-cumulative side effects.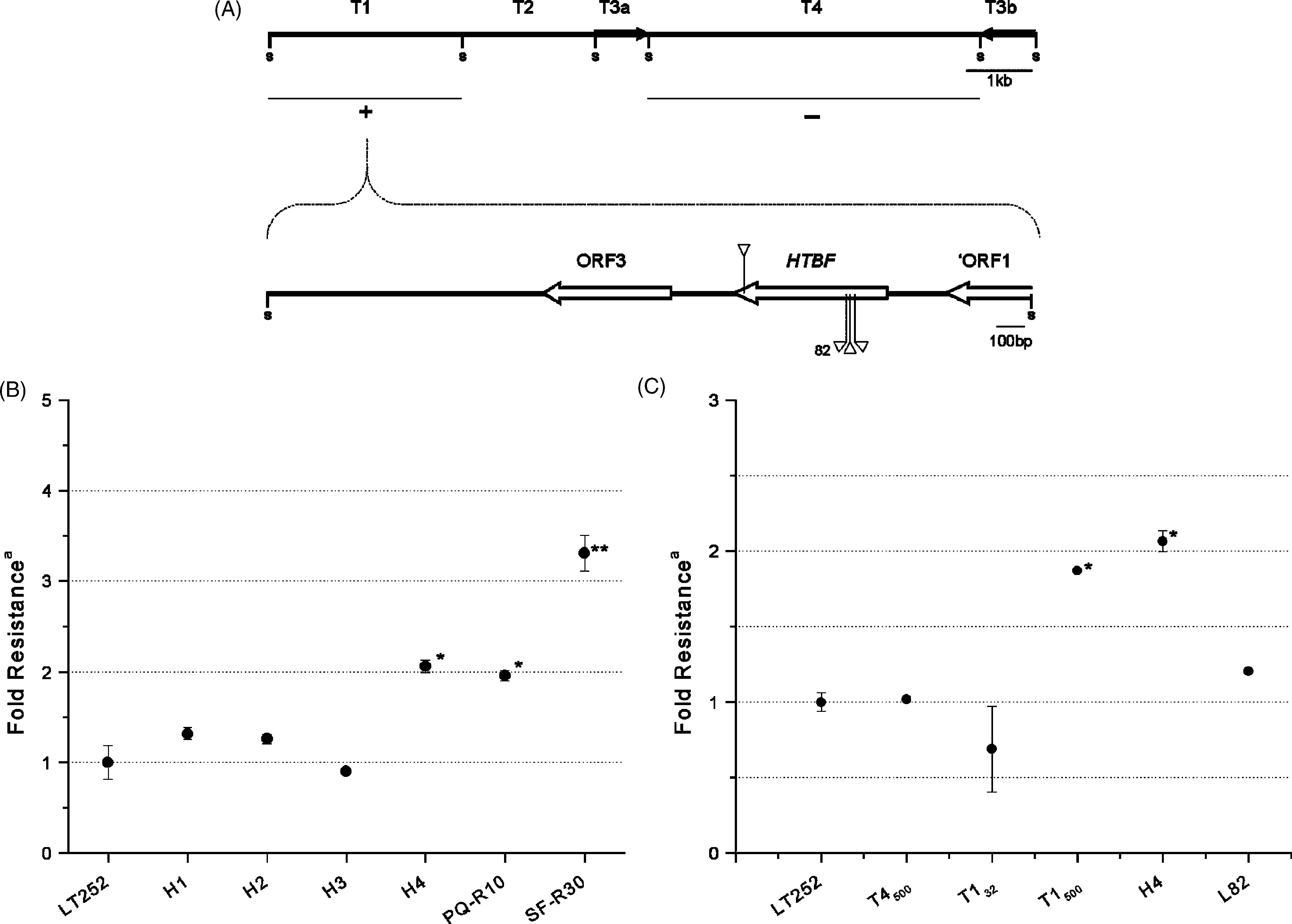 J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 1. Delimitation of the terbinafine-resistance gene within the H region. (A) SacI fragments of H4 (T1, T2, and T4) were subcloned into pELHYGand tested for terbinafine resistance; plus and minus signs represent resistance and sensitivity, respectively; black arrows represent inverted repeats (T3aand T3b); S = SacI site; sequencing of T1 revealed two complete open reading frames indicated by white arrows; the arrows on HTBF represent theplacement of mariner transposon insertion events; the insertion named 82 was used to originate the L82 transfectant. (B and C) Fold resistance of L.
J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 1. Delimitation of the terbinafine-resistance gene within the H region. (A) SacI fragments of H4 (T1, T2, and T4) were subcloned into pELHYGand tested for terbinafine resistance; plus and minus signs represent resistance and sensitivity, respectively; black arrows represent inverted repeats (T3aand T3b); S = SacI site; sequencing of T1 revealed two complete open reading frames indicated by white arrows; the arrows on HTBF represent theplacement of mariner transposon insertion events; the insertion named 82 was used to originate the L82 transfectant. (B and C) Fold resistance of L.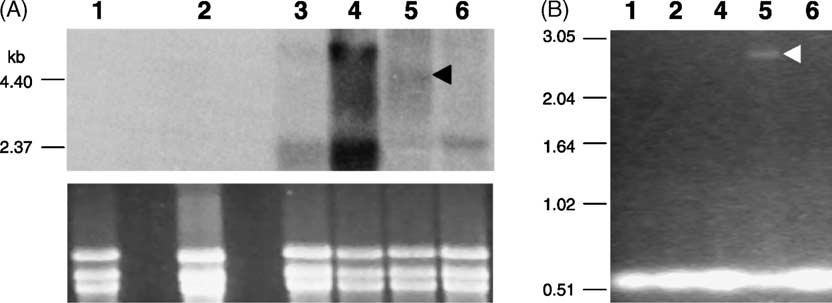 J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 2. Analyses of HTBF transcripts. (A) Northern analysis of total RNA in the cell lines LT252 (lane 1); SF-R30 (lane 6) and transfectantsT4500 (lane 2); T132 (lane 3); T1500 (lane 4) and L82 (lane 5); the black arrowhead indicates the ∼3.9 kb transcript in the transfectant bearingthe transposon insertion into HTBF. Total RNA from promastigote forms was extracted using TRIzol® reagent (Gibco BRL) and manipulated asdescribed elsewhere The HTBF probe was amplified by PCR with primers LT007 (5 -GCGCCCGGGCATATGCTCAACGAGGTGC) and LT008(5 -CGCGGATCCTAAATACCAACCAGA). (B) RT-PCR amplification products using HTBF primers (LT007/LT008) and total RNA from LT252 (lane1); T4500 (lane 2); T1500 (lane 4); L82 (lane 5); and SF-R30 (lane 6); the white arrowhead indicates the amplification product of 2.5 kb which representsthe insertion of element/GFP∗K into HTBF; reactions that did not include the reverse transcriptase confirmed the absence of contaminating DNA inthe template samples (data not shown); reverse transcriptase-PCR was performed with Ready-To-GoTM RT-PCR Beads (Amersham Pharmacia Biotech)according to the manufacturer’s specifications.
J.F.M. Marchini et al. / Molecular & Biochemical Parasitology 131 (2003) 77–81
Fig. 2. Analyses of HTBF transcripts. (A) Northern analysis of total RNA in the cell lines LT252 (lane 1); SF-R30 (lane 6) and transfectantsT4500 (lane 2); T132 (lane 3); T1500 (lane 4) and L82 (lane 5); the black arrowhead indicates the ∼3.9 kb transcript in the transfectant bearingthe transposon insertion into HTBF. Total RNA from promastigote forms was extracted using TRIzol® reagent (Gibco BRL) and manipulated asdescribed elsewhere The HTBF probe was amplified by PCR with primers LT007 (5 -GCGCCCGGGCATATGCTCAACGAGGTGC) and LT008(5 -CGCGGATCCTAAATACCAACCAGA). (B) RT-PCR amplification products using HTBF primers (LT007/LT008) and total RNA from LT252 (lane1); T4500 (lane 2); T1500 (lane 4); L82 (lane 5); and SF-R30 (lane 6); the white arrowhead indicates the amplification product of 2.5 kb which representsthe insertion of element/GFP∗K into HTBF; reactions that did not include the reverse transcriptase confirmed the absence of contaminating DNA inthe template samples (data not shown); reverse transcriptase-PCR was performed with Ready-To-GoTM RT-PCR Beads (Amersham Pharmacia Biotech)according to the manufacturer’s specifications.