Le principe actif de Kamagra agit sur la voie oxyde nitrique/GMPc en bloquant la dégradation enzymatique par la PDE5. Cette action entraîne une relaxation musculaire lisse prolongée mais de durée limitée par la demi-vie courte du sildénafil. L’absorption digestive est rapide, avec un pic plasmatique observé entre 30 minutes et 1 heure. Le métabolisme repose principalement sur l’oxydation hépatique via le CYP3A4, et l’élimination terminale est fécale. Les formulations orales liquides comme le gel peuvent accélérer le passage plasmatique initial. Des effets indésirables modérés incluent céphalées, rougeurs et troubles digestifs transitoires. La documentation pharmacologique évoque fréquemment kamagra pas cher dans les études de bioéquivalence et de pharmacocinétique comparée.
Daxitrol white paper
Daxitrol: A Novel Approach for Controlling Cravings
David Marc, B.Sc.a, & Kelly Olson Ph.Da. a NeuroScience, Inc., 373 280th St., Osceola, WI 54020, United States [email protected]ABSTRACT
Cravings are defined as an intense drive or desire directed towards food, drugs, gambling, and other stimuli. The pleasure/reward and
motivational pathways associated with cravings involve multiple neurotransmitters including dopamine, glutamate, serotonin, acetylcholine, and
various peptides. These neurotransmitters also serve as therapeutic targets to modulate the pleasure/reward pathway. Daxitrol is a novel product
that contains L-3,4-dihydroxyphenylalanine (L-DOPA), epigallocatechin gallate (EGCG), N-acetylcysteine (NAC), 5-hydroxytryptophan (5-
HTP), L-phenylalanine, forskolin, and huperzine A (HupA) to manage cravings and enhance self-control. This overview will present the
neurological pathways that underlie cravings and propose the use of Daxitrol to ameliorate cravings. Information will be provided that discusses
how Daxitrol targets dopamine, glutamate, serotonin, acetylcholine, and the peptide cholecystokinin (CCK) to help decrease cravings and
INTRODUCTION
1989). Although cravings decrease with the intake of sweets, the
Cravings are defined as an intense drive or desire for a
likelihood of weight issues increases due to their high caloric value
variety of stimuli (Pelchat, 2002). Commonly, cravings are
directed towards foods or drugs and have been reported to occur in
Whether cravings are directed towards foods or drugs,
approximately 52–97% of individuals (Gendall et al., 1997;
it is related to the lack of impulse control (Pelchat, 2002).
Christensen & Pettijohn, 2001). Cravings can also be directed
Similarly, impulsivity is observed with cravings toward other
towards behaviors that elicit a desire for intangible stimuli as
stimuli such as gambling (Iancu et al., 2008). Because cravings are
observed in gambling (Tavares et al., 2005). Interestingly, the
related to the pursuit of pleasure and reward and the impulsive
same biochemical pathways and neurological circuits are
need for a specific stimulus, cravings share similar biological
implicated in cravings for food, drugs, and gambling (Pelchat,
systems independent of the stimulus being craved (Pelchat, 2002).
2002). Daxitrol is a product that targets these neurological circuits
Therefore, a product, such as Daxitrol, that targets the biochemical
by supporting specific neurotransmitters involved in the
pathways involved with pleasure, reward, and impulsivity may
pleasure/reward pathway and therefore can decrease cravings and
reduce cravings for a variety of stimuli.
Although the biochemical mechanisms of cravings are
Food cravings are of interest because of their
not fully understood, research strongly suggests the involvement of
overwhelming prevalence and the significant impact on weight and
dopaminergic brain nuclei in regulating pleasure and reward
health. However, there is an evolutionary concept that suggests the
(Christensen & Pettijohn, 2001; Heinz et al., 2009). In addition,
purpose of food cravings is to promote greater dietary variety.
research supports the involvement of other biochemical mediators,
Evidence to support this concept has shown that adults who
such as serotonin, glutamate, acetylcholine, and various peptides in
consumed a single food diet experienced increased food cravings
behaviors related to cravings (Heinz et al., 2009). Novel strategies
as compared to a baseline diet with greater diversity (Pelchat &
are being developed to control cravings by targeting these specific
Schaefer, 2000). However, food cravings tend to be directed
biochemical pathways. This overview introduces and explains the
towards highly-palatable, calorie-dense foods such as chocolate,
neurological pathways involved in cravings and provide evidence
candy, and pizza (Weingarten & Elston, 1991). Consequently,
for the use of the novel product, Daxitrol, to manage these
cravings can contribute to weight gain (Krebs-Smith et al., 1987;
Drug and food cravings involve activation of similar
THE BIOLOGY OF CRAVINGS
biochemical pathways leading to behaviors related to an impulsive
Various neurological systems contribute to the
drive to consume a particular substance. Interestingly, programs to
behavioral drive for pleasure and reward that can lead to the onset
reduce substance use often utilize sweets as a means to reduce drug
of cravings. Of primary focus are the neurotransmitters dopamine,
cravings (Pelchat, 2002). The ingestion of sweets has shown to
glutamate, acetylcholine, and serotonin as they have been shown to
reduce cravings for opiates, alcohol, and cigarettes (Morabia et al.,
be involved in these processes (Ciccocioppo, 1999; Clay et al.,
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease.
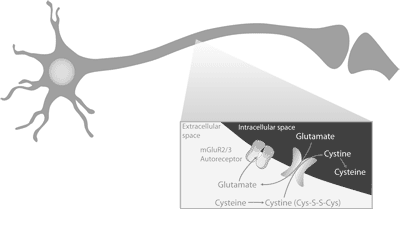
2008). In addition, a peptide called cholecystokinin (CCK) can
for intracellular glutamate, which serves as the primary source of
signal satiety, or the feeling of “fullness”, and can decrease drug
extracellular glutamate in the NAc (Knackstedt et al., 2009).
intake by altering specific actions in the central nervous system
Extracellular glutamate differs from synaptic glutamate by acting
(CNS) (Dockray, 2009; Watkins et al., 1985).
primarily on autoreceptors found on the presynaptic glutamate
Dopamine is important in reward and motivation. The
neuron, whereas, synaptic glutamate primarily acts on receptors
dopaminergic brain regions that are associated with cravings
found on the postsynaptic neuron. The normal physiological role of
include the nucleus accumbens (NAc), ventral tegmental area
extracellular glutamate in the NAc is part of a negative feedback
(VTA), and neostriatum (Clay et al., 2008). A variety of drugs
loop to prevent excessive glutamate release and reduce synaptic
such as opiods, alcohol, cocaine, and amphetamines elicit intense
glutamate levels (Figure 1; Moran et al., 2005). Research has
dopaminergic signals in the brain that induce a pleasurable
demonstrated that overtime drug use can decrease xc- transport
behavioral response (Di & Imperato, 1988). Likewise, gambling
function resulting in decreased extracellular glutamate and
can increase the activity of the same dopaminergic pathways
increased synaptic glutamate release (LaRowe et al, 2006).
(Iancu et al., 2008). Food is another powerful stimulator of
Decreased activity of the xc- transport system contributes to
dopaminergic reward regions in the brain (Epstein & Leddy,
increased cravings and perpetuation of drug use (LaRowe et al.,
2006). Upon initial exposure to a rewarding stimulus, there is an
2006; Moran et al., 2005). Decreased xc- transport function has
increase in dopamine signaling from the VTA to the NAc which
also been observed in gambling (Grant et al., 2007) and nail-biting
can result in a pleasurable response (Roberts & Koob, 1997). In
(Berk et al., 2009) as well as other craving-associated behaviors.
addition, the NAc has dense acetylcholine projections that can
New strategies which minimize alterations in the xc- transport
mediate dopamine release and enhance the reinforcing effects
system and decrease cravings are currently in development
following the ingestion of foods and drugs (Hikida et al, 2003). In
fact, research suggests that increases in brain acetylcholine can
Figure 1.
prevent reward seeking behaviors and cravings for food and drugs
Recent evidence has suggested that overeating may be a
compensatory action in response to a reward deficiency, resulting
from hypoactive dopaminergic activity in the brain (Reinholz et
al., 2008). Various conditions such as genetic predisposition or
downregulation due to preceding dopaminergic overstimulation
can lead to deficient dopaminergic stores (Reinholz et al., 2008).
Consequently, dopaminergic support may prevent compensatory
Dopamine's role in motivation is mediated by
The xc- transport system exchanges intracellular glutamate for
learned/associative processes in the brain by which exposure to
extracellular cystine. Extracellular glutamate can bind to
environmental cues can induce a craving. To explain further, drugs
presynaptic group II metabotropic autoreceptors (mGluR2/3), which inhibits synaptic glutamate signaling.
and foods increase dopamine activity in the NAc which enhances
glutamatergic signaling to the prefrontal cortex thereby creating an
While dopamine is part of the reward process that
association between the environment and the stimuli's pleasurable
triggers cravings, serotonin determines the level of motivation to
action (Goldstein & Volkow, 2002). Upon later exposure to the
satisfy the craving. Specifically, studies have reported that
environmental cues, glutamate signals from the prefrontal cortex
serotonin depletion can contribute to poor impulse control.
project to the NAc to determine the behaviors necessary to obtain
Animals that are depleted of serotonin have increased preference
the rewarding stimuli (Volkow et al., 2002). The motivation to
for a small immediate reward over a larger delayed reward (Bizot
seek and obtain these rewarding stimuli results in the manifestation
of cravings (Goldstein & Volkow, 2002).
The dopamine- and serotonin-based model has been
The glutamate signaling that promotes compulsive and
proposed as a two-stage process that triggers cravings
urge-driven behaviors is regulated by a cystine/glutamate
(Ciccocioppo, 1999). In the first stage, dopamine is released as part
antiporter, also known as the xc- transport system (Knackstedt et
of the learned processes that associate a stimulus (i.e., food, drugs,
al., 2009). The xc- transport system exchanges extracellular cystine
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease. Individual results may vary.
or gambling) with a pleasurable response and triggers the desire to
ingredient found in Daxitrol will be described in the subsequent
repeat the stimulus, thereby creating cravings. However, adequate
section to highlight how each ingredient can decrease cravings.
levels of serotonin are able to control the inclination to satisfy the
craving (Ciccocioppo, 1999; Weiss et al., 2001). In the second
A CLOSER LOOK AT DAXITROL FOR CRAVINGS
stage of the process, continued use of the stimulus results in a
Daxitrol is a dietary supplement that contains L-3,4-
depletion of serotonin and an inability to control or put-off the
desire for the stimulus (Ciccocioppo, 1999). Interestingly, many
(EGCG), N-acetylcysteine (NAC), 5-hydroxytryptophan (5-HTP),
cravings are often associated with mood issues which can also be
L-phenylalanine, forskolin, and huperzine A (HupA) (Table 1).
triggered by serotonin deficiencies (Young & Wohl, 2009;
The following overview will present how the biological
Nakama et al., 2008; Quello et al., 2005; Parker & Crawford,
mechanisms of each ingredient found in Daxitrol can help to
2007). As such, support programs designed to minimize the
neurotransmitter impediments that result in cravings can also have
L-3,4-dihydroxyphenylalanine (L-DOPA) is an amino
acid precursor to the catecholamines dopamine, norepinephrine,
An emerging area of research is now examining the role
and epinephrine (Mena et al., 2009). Low dopamine has been
of peptides in food and drug cravings. Many peptides, such as
associated with cravings to foods and drugs (Reinholz et al., 2008).
gastrin, CCK, secretin, and motilin, are found in the
Food consumption and substance use compensate for the
gastrointestinal system and can act as neuromodulators that send
hypoactive dopaminergic activity and reward deficiency (Reinholz
hunger or satiety signals to the brain (Dockray, 2009). These
et al., 2008). Ingredients that increase dopamine (Balcioglu &
peptides transmit satiety signals through cholinergic stimulation of
Wurtman, 1998), such as the L-DOPA found in Daxitrol, may
the vagus nerve to relay information from the gastrointestinal
prevent a reward deficiency and therefore decrease food and drug
system to the brain (Bodenlos et al., 2007a; Bodenlos et al.,
cravings (Reinholz et al., 2008; Palhagen et al., 2005).
2007b). Extensive research has focused on the peptide CCK and its
Epigallocatechin gallate (EGCG) acts as the primary
role in controlling food intake and cravings (Crespi et al., 2000;
polyphenolic catechin agent in green tea which is responsible for
Hirschberg et al., 2004). Interestingly, CCK is found not only in
many of green tea’s health benefits (Nagle et al., 2006). EGCG has
the gastrointestinal system but also in the brain to further regulate
antioxidant properties, promotes healthy cardiovascular function,
hunger or satiety signals (Beinfeld et al., 2002). Research has
and limits tissue damage following prolonged immune system
shown that cocaine and other drugs can lead to decreased hunger
activation (Nagle et al., 2006). Energy expenditure can also be
and food intake by increasing CCK activity in the brain (Abraham
stimulated through the thermogenic effects of EGCG (Hursel et al.,
et al., 2009, Maletinska et al., 2008). In addition, research has
2009). The thermogenic property of EGCG is attributed to its
demonstrated that brain CCK is co-released with dopamine in the
ability to inhibit catechol O-methyltransferase (COMT), thereby,
NAc where CCK may be associated with pleasure and reward
increasing the levels of dopamine and norepinephrine and cyclic
responses (Beinfeld et al., 2002). Novel strategies are in
adenosine monophosphate (cAMP) (Hursel et al., 2009). Daxitrol
development to support CCK to reduce hunger and support the
contains EGCG to support catecholamine levels and decrease
pleasure reward pathways (Ballinger & Clark, 1994; Dockray,
cravings (Sinha et al., 2003), and may aid in weight management
by enhancing thermogenesis (Hursel & Westerterp-Plantenga,
Cravings can occur via the actions of several biological
pathways. Dopamine mediates the pleasure and rewarding
N-acetylcysteine is a known precursor for cysteine
responses of stimuli whereas serotonin mediates the motivation
which is necessary for the synthesis of glutathione, a powerful
and impulsive drive to seek-out the stimuli. Acetylcholine
antioxidant (Dilger & Baker, 2007; Diniz et al., 2006). However,
enhances the dopamine activity that supports pleasure and reward.
researchers have also discovered that n-acetylcysteine possesses
Glutamate alterations can contribute to impulse behaviors and
the ability to decrease drug cravings through alterations of the xc-
cravings. Finally, CCK and acetylcholine can also contribute to
transport system (LaRowe et al., 2006; Moran et al., 2005). As
the intake of a substance by altering hunger and satiety signals.
described above, prolonged drug use can decrease xc- transport
Overall, Daxitrol is a novel dietary supplement that has been
function and lead to elevated synaptic glutamate levels that
developed to specifically support these pathways to help decrease
contribute to increased cravings (Knackstedt et al., 2009; Moran et
the prevalence of cravings. The biological actions of each
al., 2005). Oral delivery of n-acetylcysteine can reestablish normal
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease. Individual results may vary.
xc- transport function and thereby decrease reward-seeking
important in many biological processes (Henderson et al., 2005).
behavior (LaRowe et al., 2006). Altered xc- transport function is
Forskolin has been found to support neuronal catecholamine
further implicated in gambling (Grant et al., 2007) and nail-biting
activity by increasing the cAMP-accumulating properties
(Berk et al., 2009) whereby n-acetylcysteine has been reported to
following the activation of catecholamine receptors (Hoffstedt et
attenuate these behaviors. N-acetylcysteine is included in Daxitrol
al., 2001; Lang et al., 2003). Additionally, forskolin has been
to control synaptic glutamate levels and decrease impulsivity and
studied extensively for its weight management properties. Fat cells
utilize cAMP to breakdown stored fats, regulate the body’s
Daxitrol also contains 5-HTP, an amino acid precursor
thermogenic response to foods, and increase the body’s metabolic
to the monoamine neurotransmitter serotonin. Animal studies have
rate (Henderson et al., 2005). Forskolin can support weight
reported that 5-HTP reduces food intake (Amer et al., 2004) and
management by increasing cAMP which leads to the activation of
decreases impulsivity (Ho et al., 1998). 5-HTP is thought to
protein kinase A which then activates the production of hormone-
modulate these behaviors by increasing the level of serotonin in the
sensitive lipase (HSL) (Godard et al., 2005). HSL is the rate-
brain and periphery (Ho et al., 1998). Additionally, human studies
limiting step in the breakdown of triacylglycerol molecules,
have examined the use of 5-HTP and decreased cravings. A short
thereby freeing fatty acids for energy (Lafontan & Langin, 2009).
term six-week study showed that 5-HTP can reduce food intake in
Therefore, forskolin has actions that can promote favorable
overweight individuals regardless of dietary restriction (Cangiano
changes in body composition (Godard et al., 2005). The inclusion
et al., 1992). Similarly, a two-week study of overweight
of forskolin in Daxitrol could support dopamine, norepinephrine,
individuals reported reduced food intake following 5-HTP
and epinephrine activity to control cravings and assist in weight
administration compared to placebo (Cangiano et al., 1998).
Interestingly, this study also provided evidence that 5-HTP is able
Lastly, Daxitrol contains huperzine A (HupA), which is
to suppress carbohydrate and fat intake (Cangiano et al., 1998).
isolated from the Chinese herb Huperzia serrata. HupA is a potent,
5-HTP can also decrease cravings associated with drug-
highly specific, and reversible acetylcholinesterase (AChE)
use. A study in rats showed that 5-HTP can decrease the
inhibitor (Tang et al., 1989). AChE exists in multiple forms and
consumption of alcohol while increasing alcohol avoidance (Zabik
acts as the primary enzyme that breaks down acetylcholine
et al., 1994). Another study revealed that 5-HTP reduced the desire
(Brimijoin, 1983). HupA appears to preferentially inhibit the
for cocaine following cocaine withdrawal (Harris et al., 2001).
tetrameric G4 form of the acetylcholinesterase enzyme, which is
Overall, serotonin supporting agents, such as 5-HTP, can be
primarily localized in the central nervous system (Zhao & Tang,
helpful for impulse control and limit the use of craved stimuli.
2002). Therefore, HupA selectively increases acetylcholine in the
Daxitrol contains 5-HTP to increase self-control, thereby
brain (Vigny et al., 1979; Grassi et al., 1982). Research shows that
decreasing the use of craved substances.
acetylcholine is involved in the pleasure/reward pathway by
The essential amino acid, L-phenylalanine, is also
supporting the release of dopamine in the VTA and NAc (Yang et
found in Daxitrol. L-phenylalanine can be beneficial for mood and
al., 2009; Shirayama & Chaki, 2006). It has been proposed that
focus by supporting the synthesis of L-tyrosine (a precursor to
cholinesterase inhibitors decrease cravings by increasing
dopamine, norepinephrine, and epinephrine) and phenylethylamine
acetylcholine in the brain, thereby promoting the binding of the
(PEA) (Kusaga et al., 2002). In addition, L-phenylalanine can
nicotinic acetylcholine receptor (Alisky, 2006). The binding of
enhance satiety in humans by inducing the release of CCK
acetylcholine on the nicotinic acetylcholine receptor can promote
(Ballinger & Clark, 1994; Pohle-Krauza et al., 2008), although the
dopamine activity and increase pleasurable responses, ultimately,
precise mechanism is not completely understood. Increased CCK
decreasing the need for rewarding stimuli (Alisky, 2006; Hiranita
levels can decrease food intake by enhancing satiety signals that
are transmitted from the gastrointestinal system to the brain
The vagus nerve also utilizes acetylcholine as its
(Ballinger & Clark, 1994). Therefore the inclusion of L-
primary chemical messenger and provides a source of
phenylalanine in Daxitrol helps to promote the feeling of
acetylcholine in the brain (Bodenlos et al., 2007a; Bodenlos et al.,
“fullness” and limit the opportunity to develop cravings.
2007b). As mentioned above, the vagus nerve mediates
Forskolin, isolated from the plant Coleus forskohli, can
communication between the gastrointestinal system and the brain
directly increase the activity of adenylate cyclase (Burns et al.,
to signal satiety (Bodenlos et al., 2007a; Bodenlos et al., 2007b).
1987). Adenylate cyclase is an enzyme that activates cyclic
Therefore, HupA may help facilitate and promote satiety signals in
adenosine monophosphate (cAMP), which is a second messenger
addition to increasing pleasurable responses to decrease food and
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease. Individual results may vary.
drug intake. Daxitrol includes HupA as a way to support
Bodenlos, J. S., Kose, S., Borckardt, J. J., Nahas, Z., Shaw, O'Neil, P. M. et al. (2007a). Vagus
Nerve Stimulation and Emotional Responses to Food among Depressed Patients.
acetylcholine function in the brain, which may assist in decreasing
J.Diabetes Sci.Technol., 1, 771-779.
Bodenlos, J. S., Kose, S., Borckardt, J. J., Nahas, Z., Shaw, D., O'Neil, P. M. et al. (2007b).
Vagus nerve stimulation acutely alters food craving in adults with depression. Appetite, 48, 145-153.
Brimijoin, S. (1983). Molecular forms of acetylcholinesterase in brain, nerve and muscle: nature,
localization and dynamics. Prog.Neurobiol., 21, 291-322.
Burns, T. W., Langley, P. E., Terry, B. E., Bylund, D. B., & Forte, L. R., Jr. (1987). Comparative
effects of forskolin and isoproterenol on the cyclic AMP content of human adipocytes. Life Sci., 40, 145-154.
Ingredient Neurological Targets Reference
Cangiano, C., Ceci, F., Cascino, A., Del, B. M., Laviano, A., Muscaritoli, M. et al. (1992). Eating
behavior and adherence to dietary prescriptions in obese adult subjects treated with 5-hydroxytryptophan. Am.J.Clin.Nutr., 56, 863-867.
Cangiano, C., Laviano, A., Del, B. M., Preziosa, I., Angelico, F., Cascino, A. et al. (1998).
Effects of oral 5-hydroxy-tryptophan on energy intake and macronutrient
selection in non-insulin dependent diabetic patients. Int.J.Obes.Relat Metab Disord., 22, 648-654.
Christensen, L. & Pettijohn, L. (2001). Mood and carbohydrate cravings. Appetite, 36, 137-145.
Ciccocioppo, R. (1999). The role of serotonin in craving: from basic research to human studies.
Clay, S. W., Allen, J., & Parran, T. (2008). A review of addiction. Postgrad.Med., 120, E01-E07.
Crespi, F., Corsi, M., Reggiani, A., Ratti, E., & Gaviraghi, G. (2000). Involvement of
cholecystokinin within craving for cocaine: role of cholecystokinin receptor ligands. Expert.Opin.Investig.Drugs, 9, 2249-2258.
CONCLUSION
Di, C. G. & Imperato, A. (1988). Drugs abused by humans preferentially increase synaptic
Cravings can be associated with weight issues, drug
dopamine concentrations in the mesolimbic system of freely moving rats. Proc.Natl.Acad.Sci.U.S.A, 85, 5274-5278.
use, gambling, and other behaviors. The purpose of Daxitrol is to
Dilger, R. N. & Baker, D. H. (2007). Oral N-acetyl-L-cysteine is a safe and effective precursor of
cysteine. J.Anim Sci., 85, 1712-1718.
not only decrease cravings but also provide the necessary support
Diniz, Y. S., Rocha, K. K., Souza, G. A., Galhardi, C. M., Ebaid, G. M., Rodrigues, H. G. et al.
to enhance self-control, to limit the consumption of craved stimuli
(2006). Effects of N-acetylcysteine on sucrose-rich diet-induceratsd hyperglycaemia, dyslipidemia and oxidative stress in. Eur.J.Pharmacol., 543,
and support healthy weight. The ingredients in Daxitrol decrease
cravings by supporting dopaminergic, glutamatergic, serotonergic,
Dockray, G. J. (2009). Cholecystokinin and gut-brain signalling. Regul.Pept., 155, 6-10.
Epstein, L. H. & Leddy, J. J. (2006). Food reinforcement. Appetite, 46, 22-25.
acetylcholinergic, and peptidergic systems. These biochemical
Gendall, K. A., Joyce, P. R., & Sullivan, P. F. (1997). Impact of definition on prevalence of food
systems regulate pleasure, reward, motivation, impulsivity, and
cravings in a random sample of young women. Appetite, 28, 63-72.
satiety. The commonality of all these actions is their role in
Godard, M. P., Johnson, B. A., & Richmond, S. R. (2005). Body composition and hormonal
adaptations associated with forskolin consumption in overweight and obese men.
cravings. Overall, Daxitrol is a novel and unique dietary
supplement that targets various biochemical mediators to decrease
Goldstein, R. Z. & Volkow, N. D. (2002). Drug addiction and its underlying neurobiological
basis: neuroimaging evidence for the involvement of the frontal cortex. Am.J.Psychiatry, 159, 1642-1652.
Grant, J. E., Kim, S. W., & Odlaug, B. L. (2007). N-acetyl cysteine, a glutamate-modulating
agent, in the treatment of pathological gambling: a pilot study. Biol.Psychiatry, 62, 652-657.
REFERENCES
Grassi, J., Vigny, M., & Massoulie, J. (1982). Molecular forms of acetylcholinesterase in bovine
caudate nucleus and superior cervical ganglion: solubility properties and
Abraham, H., Covasa, M., & Hajnal, A. (2009). Cocaine- and amphetamine-regulated transcript
hydrophobic character. J.Neurochem., 38, 457-469.
peptide immunoreactivity in the brain of the CCK-1 receptor deficient obese OLETF rat. Exp.Brain Res., 196, 545-556.
Harris, G. C., Altomare, K., & ston-Jones, G. (2001). Preference for a cocaine-associated
environment is attenuated by augmented accumbal serotonin in cocaine
Alisky, J. M. (2006). Cholinesterase inhibitors might alleviate methamphetamine-induced
withdrawn rats. Psychopharmacology (Berl), 156, 14-22.
delusions, hallucinations and cognitive impairment, while reducing craving and addiction. World J.Biol.Psychiatry, 7, 269.
Heinz, A., Beck, A., Grusser, S. M., Grace, A. A., & Wrase, J. (2009). Identifying the neural
circuitry of alcohol craving and relapse vulnerability. Addict.Biol., 14, 108-118.
Amer, A., Breu, J., McDermott, J., Wurtman, R. J., & Maher, T. J. (2004). 5-Hydroxy-L-
tryptophan suppresses food intake in food-deprived and stressed rats.
Henderson, S., Magu, B., Rasmussen, C., Lancaster, S., Kerksick, C., Smith, P. et al. (2005).
Pharmacol.Biochem.Behav., 77, 137-143.
Effects of coleus forskohlii supplementation on body composition and hematological profiles in mildly overweight women. J.Int.Soc.Sports Nutr., 2,
Balcioglu, A. & Wurtman, R. J. (1998). Effects of fenfluramine and phentermine (fen-phen) on
dopamine and serotonin release in rat striatum: in vivo microdialysis study in conscious animals. Brain Res., 813, 67-72.
Hikida, T., Kitabatake, Y., Pastan, I., & Nakanishi, S. (2003). Acetylcholine enhancement in the
nucleus accumbens prevents addictive behaviors of cocaine and morphine.
Ballinger, A. B. & Clark, M. L. (1994). L-phenylalanine releases cholecystokinin (CCK) and is
associated with reduced food intake in humans: evidence for a physiological role of CCK in control of eating. Metabolism, 43, 735-738.
Hiranita, T., Nawata, Y., Sakimura, K., Anggadiredja, K., & Yamamoto, T. (2006). Suppression
of methamphetamine-seeking behavior by nicotinic agonists.
Beinfeld, M. C., Connolly, K. J., & Pierce, R. C. (2002). Cocaine treatment increases
Proc.Natl.Acad.Sci.U.S.A, 103, 8523-8527.
extracellular cholecystokinin (CCK) in the nucleus accumbens shell of awake, freely moving rats, an effect that is enhanced in rats that are behaviorally
Hirschberg, A. L., Naessen, S., Stridsberg, M., Bystrom, B., & Holtet, J. (2004). Impaired
sensitized to cocaine. J.Neurochem., 81, 1021-1027.
cholecystokinin secretion and disturbed appetite regulation in women with polycystic ovary syndrome. Gynecol.Endocrinol., 19, 79-87.
Berk, M., Jeavons, S., Dean, O. M., Dodd, S., Moss, K., Gama, C. S. et al. (2009). Nail-biting
stuff? The effect of N-acetyl cysteine on nail-biting. CNS.Spectr., 14, 357-360.
Ho, M. Y., Al-Zahrani, S. S., Al-Ruwaitea, A. S., Bradshaw, C. M., & Szabadi, E. (1998). 5-
hydroxytryptamine and impulse control: prospects for a behavioural analysis.
Billes, S. K. & Cowley, M. A. (2008). Catecholamine reuptake inhibition causes weight loss by
increasing locomotor activity and thermogenesis. Neuropsychopharmacology, 33, 1287-1297.
Hoffstedt, J., Arner, P., Schalling, M., Pedersen, N. L., Sengul, S., Ahlberg, S. et al. (2001). A
common hormone-sensitive lipase i6 gene polymorphism is associated with
Bizot, J., Le, B. C., Puech, A. J., Hamon, M., & Thiebot, M. (1999). Serotonin and tolerance to
decreased human adipocyte lipolytic function. Diabetes, 50, 2410-2413.
delay of reward in rats. Psychopharmacology (Berl), 146, 400-412.
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease. Individual results may vary.
Hursel, R., Viechtbauer, W., & Westerterp-Plantenga, M. S. (2009). The effects of green tea on
Vigny, M., Bon, S., Massoulie, J., & Gisiger, V. (1979). The subunit structure of mammalian
weight loss and weight maintenance: a meta-analysis. Int.J.Obes.(Lond).
acetylcholinesterase: catalytic subunits, dissociating effect of proteolysis and disulphide reduction on the polymeric forms. J.Neurochem., 33, 559-562.
Hursel, R. & Westerterp-Plantenga, M. S. (2009). Green tea catechin plus caffeine
supplementation to a high-protein diet has no additional effect on body weight
Volkow, N. D., Fowler, J. S., Wang, G. J., & Goldstein, R. Z. (2002). Role of dopamine, the
maintenance after weight loss. Am.J.Clin.Nutr., 89, 822-830.
frontal cortex and memory circuits in drug addiction: insight from imaging studies. Neurobiol.Learn.Mem., 78, 610-624.
Iancu, I., Lowengrub, K., Dembinsky, Y., Kotler, M., & Dannon, P. N. (2008). Pathological
gambling: an update on neuropathophysiology and pharmacotherapy.
Watkins, L. R., Kinscheck, I. B., Kaufman, E. F., Miller, J., Frenk, H., & Mayer, D. J. (1985).
Cholecystokinin antagonists selectively potentiate analgesia induced by endogenous opiates. Brain Res., 327, 181-190.
Knackstedt, L. A., LaRowe, S., Mardikian, P., Malcolm, R., Upadhyaya, H., Hedden, S. et al.
(2009). The role of cystine-glutamate exchange in nicotine dependence in rats
Weingarten, H. P. & Elston, D. (1991). Food cravings in a college population. Appetite, 17, 167-
and humans. Biol.Psychiatry, 65, 841-845.
Krebs-Smith, S. M., Smiciklas-Wright, H., Guthrie, H. A., & Krebs-Smith, J. (1987). The effects
Weiss, F., Ciccocioppo, R., Parsons, L. H., Katner, S., Liu, X., Zorrilla, E. P. et al. (2001).
of variety in food choices on dietary quality. J.Am.Diet.Assoc., 87, 897-903.
Compulsive drug-seeking behavior and relapse. Neuroadaptation, stress, and conditioning factors. Ann.N.Y.Acad.Sci., 937, 1-26.
Kusaga A, Yamashita Y, Koeda T, et al (2002). Increased urine phenylethylamine after
methylphenidate treatment in children with ADHD. Ann. Neurol., 52, 372-374.
Yang, K., Hu, J., Lucero, L., Liu, Q., Zheng, C., Zhen, X. et al. (2009). Distinctive nicotinic
acetylcholine receptor functional phenotypes of rat ventral tegmental area
Lafontan, M. & Langin, D. (2009). Lipolysis and lipid mobilization in human adipose tissue.
dopaminergic neurons. J.Physiol, 587, 345-361.
Young, M. M. & Wohl, M. J. (2009). "The Gambling Craving Scale: Psychometric validation and
Lang, J. H., Zhu, L., Sun, Z. J., & Chen, J. (2003). Estrogen levels and estrogen receptors in
behavioral outcomes": Correction to Young and Wohl (2009).
patients with stress urinary incontinence and pelvic organ prolapse.
Int.J.Gynaecol.Obstet., 80, 35-39.
Zabik, J. E., Sprague, J. E., & Binkerd, K. (1994). Central and peripheral components of the
LaRowe, S. D., Mardikian, P., Malcolm, R., Myrick, H., Kalivas, P., McFarland, K. et al. (2006).
inhibitory actions of 5-HTP on ethanol consumption in the rat.
Safety and tolerability of N-acetylcysteine in cocaine-dependent individuals.
Pharmacol.Biochem.Behav., 47, 547-551.
Zhao, Q. & Tang, X. C. (2002). Effects of huperzine A on acetylcholinesterase isoforms in vitro:
Maletinska, L., Maixnerova, J., Matyskova, R., Haugvicova, R., Pirnik, Z., Kiss, A. et al. (2008).
comparison with tacrine, donepezil, rivastigmine and physostigmine.
Synergistic effect of CART (cocaine- and amphetamine-regulated transcript)
peptide and cholecystokinin on food intake regulation in lean mice. BMC.Neurosci., 9, 101.
Martin, C. K., O'Neil, P. M., Tollefson, G., Greenway, F. L., & White, M. A. (2008). The
association between food cravings and consumption of specific foods in a
laboratory taste test. Appetite, 51, 324-326.
Mena, M. A., Casarejos, M. J., Solano, R. M., & de Yebenes, J. G. (2009). Half a century of L-
DOPA. Curr.Top.Med.Chem., 9, 880-893.
Morabia, A., Fabre, J., Chee, E., Zeger, S., Orsat, E., & Robert, A. (1989). Diet and opiate
addiction: a quantitative assessment of the diet of non-institutionalized opiate addicts. Br.J.Addict., 84, 173-180.
Moran, M. M., McFarland, K., Melendez, R. I., Kalivas, P. W., & Seamans, J. K. (2005).
Cystine/glutamate exchange regulates metabotropic glutamate receptor presynaptic inhibition of excitatory transmission and vulnerability to cocaine seeking. J.Neurosci., 25, 6389-6393.
Nagle, D. G., Ferreira, D., & Zhou, Y. D. (2006). Epigallocatechin-3-gallate (EGCG): chemical
and biomedical perspectives. Phytochemistry, 67, 1849-1855.
Nakama, H., Chang, L., Cloak, C., Jiang, C., Alicata, D., & Haning, W. (2008). Association
between psychiatric symptoms and craving in methamphetamine users. Am.J.Addict., 17, 441-446.
Palhagen, S., Lorefalt, B., Carlsson, M., Ganowiak, W., Toss, G., Unosson, M. et al. (2005). Does
L-dopa treatment contribute to reduction in body weight in elderly patients with Parkinson's disease? Acta Neurol.Scand., 111, 12-20.
Parker, G. & Crawford, J. (2007). Chocolate craving when depressed: a personality marker.
Pelchat, M. L. (2002). Of human bondage: food craving, obsession, compulsion, and addiction.
Pelchat, M. L. & Schaefer, S. (2000). Dietary monotony and food cravings in young and elderly
adults. Physiol Behav., 68, 353-359.
Pohle-Krauza, R. J., Navia, J. L., Madore, E. Y., Nyrop, J. E., & Pelkman, C. L. (2008). Effects
of L-phenylalanine on energy intake in overweight and obese women: interactions with dietary restraint status. Appetite, 51, 111-119.
Quello, S. B., Brady, K. T., & Sonne, S. C. (2005). Mood disorders and substance use disorder: a
complex comorbidity. Sci.Pract.Perspect., 3, 13-21.
Reinholz, J., Skopp, O., Breitenstein, C., Bohr, I., Winterhoff, H., & Knecht, S. (2008).
Compensatory weight gain due to dopaminergic hypofunction: new evidence and own incidental observations. Nutr.Metab (Lond), 5, 35.
Roberts, A. J. & Koob, G. F. (1997). The neurobiology of addiction: an overview. Alcohol Health
Shirayama, Y. & Chaki, S. (2006). Neurochemistry of the nucleus accumbens and its relevance to
depression and antidepressant action in rodents. Curr.Neuropharmacol., 4, 277-291.
Sinha, R., Talih, M., Malison, R., Cooney, N., Anderson, G. M., & Kreek, M. J. (2003).
Hypothalamic-pituitary-adrenal axis and sympatho-adreno-medullary responses during stress-induced and drug cue-induced cocaine craving states. Psychopharmacology (Berl), 170, 62-72.
Tang, X. C., De, S. P., Sugaya, K., & Giacobini, E. (1989). Effect of huperzine A, a new
cholinesterase inhibitor, on the central cholinergic system of the rat. J.Neurosci.Res., 24, 276-285.
Tavares, H., Zilberman, M. L., Hodgins, D. C., & el-Guebaly, N. (2005). Comparison of craving
between pathological gamblers and alcoholics. Alcohol Clin.Exp.Res., 29, 1427-1431.
*These statements have not been evaluated by the Food and Drug Administration. This product is not intended to diagnose, treat, cure, or prevent any disease. Individual results may vary.
Edwin Winkel DDS PhD Actinobacillus actinomycetemcomitans , Bacteroides gingivalis and Bacteroides intermedius : predictors of attachment loss? Wennström JL, Dahlén G, Svensson Svensson J, Nyman S Oral Microbiol. Immunol 1987: 2:158-163 It appeared that the absence of these “indicator” bacteria is a better predictor of no further loss of atta
 2008). In addition, a peptide called cholecystokinin (CCK) can
for intracellular glutamate, which serves as the primary source of
signal satiety, or the feeling of “fullness”, and can decrease drug
extracellular glutamate in the NAc (Knackstedt et al., 2009).
intake by altering specific actions in the central nervous system
Extracellular glutamate differs from synaptic glutamate by acting
(CNS) (Dockray, 2009; Watkins et al., 1985).
primarily on autoreceptors found on the presynaptic glutamate
Dopamine is important in reward and motivation. The
neuron, whereas, synaptic glutamate primarily acts on receptors
dopaminergic brain regions that are associated with cravings
found on the postsynaptic neuron. The normal physiological role of
include the nucleus accumbens (NAc), ventral tegmental area
extracellular glutamate in the NAc is part of a negative feedback
(VTA), and neostriatum (Clay et al., 2008). A variety of drugs
loop to prevent excessive glutamate release and reduce synaptic
such as opiods, alcohol, cocaine, and amphetamines elicit intense
glutamate levels (Figure 1; Moran et al., 2005). Research has
dopaminergic signals in the brain that induce a pleasurable
demonstrated that overtime drug use can decrease xc- transport
behavioral response (Di & Imperato, 1988). Likewise, gambling
function resulting in decreased extracellular glutamate and
can increase the activity of the same dopaminergic pathways
increased synaptic glutamate release (LaRowe et al, 2006).
(Iancu et al., 2008). Food is another powerful stimulator of
Decreased activity of the xc- transport system contributes to
dopaminergic reward regions in the brain (Epstein & Leddy,
increased cravings and perpetuation of drug use (LaRowe et al.,
2006). Upon initial exposure to a rewarding stimulus, there is an
2006; Moran et al., 2005). Decreased xc- transport function has
increase in dopamine signaling from the VTA to the NAc which
also been observed in gambling (Grant et al., 2007) and nail-biting
can result in a pleasurable response (Roberts & Koob, 1997). In
(Berk et al., 2009) as well as other craving-associated behaviors.
addition, the NAc has dense acetylcholine projections that can
New strategies which minimize alterations in the xc- transport
mediate dopamine release and enhance the reinforcing effects
system and decrease cravings are currently in development
following the ingestion of foods and drugs (Hikida et al, 2003). In
fact, research suggests that increases in brain acetylcholine can
Figure 1.
2008). In addition, a peptide called cholecystokinin (CCK) can
for intracellular glutamate, which serves as the primary source of
signal satiety, or the feeling of “fullness”, and can decrease drug
extracellular glutamate in the NAc (Knackstedt et al., 2009).
intake by altering specific actions in the central nervous system
Extracellular glutamate differs from synaptic glutamate by acting
(CNS) (Dockray, 2009; Watkins et al., 1985).
primarily on autoreceptors found on the presynaptic glutamate
Dopamine is important in reward and motivation. The
neuron, whereas, synaptic glutamate primarily acts on receptors
dopaminergic brain regions that are associated with cravings
found on the postsynaptic neuron. The normal physiological role of
include the nucleus accumbens (NAc), ventral tegmental area
extracellular glutamate in the NAc is part of a negative feedback
(VTA), and neostriatum (Clay et al., 2008). A variety of drugs
loop to prevent excessive glutamate release and reduce synaptic
such as opiods, alcohol, cocaine, and amphetamines elicit intense
glutamate levels (Figure 1; Moran et al., 2005). Research has
dopaminergic signals in the brain that induce a pleasurable
demonstrated that overtime drug use can decrease xc- transport
behavioral response (Di & Imperato, 1988). Likewise, gambling
function resulting in decreased extracellular glutamate and
can increase the activity of the same dopaminergic pathways
increased synaptic glutamate release (LaRowe et al, 2006).
(Iancu et al., 2008). Food is another powerful stimulator of
Decreased activity of the xc- transport system contributes to
dopaminergic reward regions in the brain (Epstein & Leddy,
increased cravings and perpetuation of drug use (LaRowe et al.,
2006). Upon initial exposure to a rewarding stimulus, there is an
2006; Moran et al., 2005). Decreased xc- transport function has
increase in dopamine signaling from the VTA to the NAc which
also been observed in gambling (Grant et al., 2007) and nail-biting
can result in a pleasurable response (Roberts & Koob, 1997). In
(Berk et al., 2009) as well as other craving-associated behaviors.
addition, the NAc has dense acetylcholine projections that can
New strategies which minimize alterations in the xc- transport
mediate dopamine release and enhance the reinforcing effects
system and decrease cravings are currently in development
following the ingestion of foods and drugs (Hikida et al, 2003). In
fact, research suggests that increases in brain acetylcholine can
Figure 1.