Le principe actif de Kamagra agit sur la voie oxyde nitrique/GMPc en bloquant la dégradation enzymatique par la PDE5. Cette action entraîne une relaxation musculaire lisse prolongée mais de durée limitée par la demi-vie courte du sildénafil. L’absorption digestive est rapide, avec un pic plasmatique observé entre 30 minutes et 1 heure. Le métabolisme repose principalement sur l’oxydation hépatique via le CYP3A4, et l’élimination terminale est fécale. Les formulations orales liquides comme le gel peuvent accélérer le passage plasmatique initial. Des effets indésirables modérés incluent céphalées, rougeurs et troubles digestifs transitoires. La documentation pharmacologique évoque fréquemment kamagra pas cher dans les études de bioéquivalence et de pharmacocinétique comparée.
Doi:10.1016/j.plantsci.2004.11.00
Genetic transformation of the medicinal plant Ruta graveolens L.
by an Agrobacterium tumefaciens-mediated method
Karine Lie`Alain HehnThi Leˆ Minh TAntoine Gravo
Brigitte ThomassetFre´de´ric Bourgaud, Eric
aUMR 1121 INRA-INPL, Laboratoire Agronomie et Environnement, ENSAIA-2, Av. Foreˆt de Haye,
54505 Vandoeuvre-le`s-Nancy Cedex, France
bUMR-CNRS 6022, Ge´nie enzymatique et Cellulaire, UTC, BP 20529-60205 Compie`gne Cedex, France
Received 13 September 2004; received in revised form 20 October 2004; accepted 3 November 2004
Ruta graveolens L. is a source of pharmacologically active molecules such as furocoumarins, furoquinolines and acridone alkaloids. A
genetic transformation method using Agrobacterium tumefaciens was developed for this plant species. The conditions for an efficientregeneration of R. graveolens led us to retain hypocotyls among other explants tested. Subsequently, transformation was obtained by co-cultivation of hypocotyls of 2–3 weeks old plants and A. tumefaciens strain C58C1RifR containing a plasmid harboring neomycinphosphotransferase and b-glucuronidase encoding genes. PCR analyses using b-glucuronidase-primers showed that 78% of the transgenicplants selected for kanamycin resistance were transformed with the gene encoding b-glucuronidase and 67% of them exhibited thecorresponding enzymatic activity. Chromosomic integration of b-glucuronidase encoding gene was verified by Southern blotting. Routinetransformation efficiency of R. graveolens L. was established at 11% and could reach 22% in some experiments. This is the first report of amethod to transform genetically R. graveolens L. # 2004 Elsevier Ireland Ltd. All rights reserved.
Keywords: Agrobacterium tumefaciens; Ruta graveolens; Hypocotyl; Transgenic plant; gus; nptII
protect the tissues against UV light, insects and differentpathogens . Amongst furocoumarins, bergapten has been
Since ancient times, garden rue (Ruta graveolens L.,
used for decades for the treatment of various skin diseases
Rutaceae) has been among the key plants of the European
such as vitiligo and psoriasis . At present, little is known
pharmacopoeia Its medicinal value is due to the
about enzymes and the corresponding genes involved in the
numerous secondary metabolites it contains like furocou-
synthesis of furocoumarins in plants . These molecules
marins, furoquinolines and acridone alkaloids. In plant,
remain one of the least unknown elements of the
furocoumarins exhibit many physiological properties as they
phenylpropanoid pathway. Genetic transformation is apowerful tool to study gene functions in plants and couldhelp establishing new knowledge on the R. graveolens
Abbreviations: BAP, benzylaminopurin; IAA, indolacetic acid; CEF,
furocoumarin pathway. To date, there is no protocol
cefotaxim; KAN, kanamycin; KANR plant, plant resistant to kanamycin;
available to genetically engineer R. graveolens L. Two
Bp, base pair; w/v, weight/volume; MS, Murashige and Skoog medium [7];MS(30), MS medium supplemented with 30 g/l sucrose; MS(30)BAP0.1,
previous reports mentioned that R. graveolens was
MS(30) supplemented with 0.1 mg/l BAP; SE (selection efficiency), per-
susceptible to Agrobacterium wild strains. Eilert (1989) and
centage of selected kanamycin-resistant explants that developed transgenic
Porter (1991) reported, respectively, the regeneration of
plants; TE (transformation efficiency), percentage of initial explants that
possible shooty teratomas and hairy roots from R. grave-
regenerated at least one transgenic plant detected by PCR
olens. However, these authors neither genetically nor
* Corresponding author. Tel.: +33 3 83 59 57 81; fax: +33 3 83 59 57 99.
E-mail address: [email protected] (E. Gontier).
biochemically confirmed the status of these transformants.
0168-9452/$ – see front matter # 2004 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.plantsci.2004.11.001
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
These two preliminary papers suggested us to choose an
Cambridge, UK. The plasmid pBin-gus contains sequences
Agrobacterium-mediated method for the genetic transfor-
encoding resistance to spectinomycin, streptomycin and
mation of R. graveolens L. This paper describes the
ampicillin as well as the T-DNA. This T-DNA is composed
experiments that were carried out in order to obtain a fast
of a neomycin phosphotransferase (nptII) marker gene
and reliable method to genetically transform R. graveolens
driven by the nopaline synthase (nos) promoter, allowing
L. We successively determined (i) which explant was best
kanamycin (KAN) selection, and the b-glucuronidase (gus)
adapted to an efficient regeneration of a whole plant and
reporter gene, driven by the cauliflower mosaic virus (CaMV
(ii) which conditions to apply to genetically transform
35S promoter) allowing GUS histochemical assay.
explants with an Agrobacterium-mediated method. This is
Bacteria were grown for 3 days on Petri dishes with YEB
the first report of a method to genetically transform R.
(beef extract, 5 g/l; yeast extract, 1 g/l; peptone, 5 g/l;
sucrose, 5 g/l; MgSO4, 0.25 g/l; pH 7.2; difco-agar, 15 g/l)supplemented with 100 mg/l carbenicillin, 100 mg/l rifam-picin, 100 mg/l spectinomycin and 300 mg/l streptomycin at
28 8C. For transformation, a colony from the plate waspicked up and grown 12–16 h at 28 8C in YEB liquid
medium on a shaker. Bacterial cells were collected bycentrifugation (3500 rpm for 15 min), washed twice in
Basal medium MS(30) used for the cultures contained
lambda buffer (10 mM Tris–HCl, 10 mM MgSO4, pH 7.2),
MS nutrients and 30 g/l sucrose (pH 5.8). Unless
and then resuspended in MS(30)BAP0.1 (50 ml for 1 ml of
stated otherwise, all MS(30) media described in the
experiments were solidified with 0.8% Difco agar. Growthregulators were added to the media before autoclaving
(20 min at 120 8C, 105 Pa). Cultures were maintained in15 mm  57 mm Petri dishes or in glass vessels (70 mm
Factors affecting transformation frequency of R. grave-
 55 mm or 100 mm  55 mm, closed with magenta cap
olens L. were tested including pre-culture of explants (0, 1 or
purchased from Sigma) containing 10, 25 or 40 ml of
2 days), addition of acetosyringone to the co-culture medium
medium, respectively. All cultures were sealed with gas-
(0, 50 and 250 mM), and selection methods. Two types of
porous tape and kept in growth chambers at 22 8C and a 16 h
selection methods were tested. The one step selection
photoperiod with 160 mmol m2 sÀ1 light irradiance.
protocol consisted in selecting explants that resisted to50 mg/l KAN in the medium. The two step selection
protocol used explants cultivated for 3 days on solidifiedMS(30)BAP0.1 containing 300 mg/l CEF before being
R. graveolens L. seeds were purchased from Conrad
transferred onto fresh solidified MS(30)BAP0.1 containing
Appel (Samen und Pflanzen GmbH, Darmstadt, Germany).
They were scarified 10 min in 95% sulfuric acid, rinsed three
For each experiment, 200–250 explants were immersed
times with tap water, then sterilized for 5 min in 7% (w/v)
for 15 min in the Agrobacterium suspensions, then blotted
calcium hypochlorite, and finally rinsed three times for
with sterile filter paper and incubated for 3 days at 25 8C on
5 min in sterile distilled water before being germinated on
MS(30) medium. After 2–3 weeks, apices, hypocotyls,
After co-culture, explants were rinsed once in sterile
cotyledons, roots and root tips were used as primary explants
water for 15 min, blotted on sterile filter paper, rinsed once
to determine the best regeneration conditions for R.
again for 15 min in MS(30)BAP0.1 containing 500 mg/l
graveolens L. MS(30) medium supplemented with BAP
CEF and blotted on a new sterile filter paper. Explants
and/or IAA with concentrations ranging from 0 to 2 mg/l
were then transferred on selection medium containing
was used throughout these experiments.
CEF and KAN. When pre-culture was tested, explants werepreconditioned
The selection media were composed of 0.8% agar-
The disarmed A. tumefaciens C58C1RifR strain carrying
solidified MS(30)BAP0.1 added with CEF either alone or
the plasmids pGV2260 and the binary vector pBin-gus
with KAN. CEF and KAN concentrations used for selection
was used as a vector system for genetic transformation.
were determined after preliminary experiments revealing
The plasmid pGV2260 carries the vir genes necessary to
lethal and sub-lethal doses (data not shown).
infect the plant plus a gene encoding carbenicillin resistance.
After 4–6 weeks on selection media, developing shoots
The plasmid pBin-gus was obtained by replacing the
were excised from the explants and transferred for 3 weeks
mGFP5ER gene by the gus gene in the plasmid pBin-
into 55 mm  70 mm glass vessels containing MS(30)-
mGFP5ER obtained from Prof. Jim Haseloff, MRC
BAP0.1 with 300 mg/l CEF and 75 mg/l KAN. A maximum
Laboratory of Molecular Biology, Addenbrookes Hospital,
of one shoot was taken from each extremity of the hypocotyl
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
to ensure that all the transformed shoots were coming fromindependent transformation events.
The shoots were then transferred for 6–8 weeks in
55 mm  100 mm glass vessels containing MS(30) for rootformation. Plants were acclimated and transferred into aphytotronic chamber.
Fig. 1. Effect of BAP on direct shoot regeneration applied to different
For histochemical GUS assay, freshly harvested leaves of
explants. Apices, cotyledons, hypocotyls, roots and root tips were cultivated
entire plants were immersed into a solution of 1 mM 3-bromo-
on agar-solidified MS(30) containing different concentrations of BAP (0,
4-chloro-3-indolyl glucuronid (X-gluc) and 100 mM Tris–
0.1, 1 and 2 mg/l). Shoot regeneration was evaluated after 2 weeks for each
HCl and 50 mM NaCl, pH 7 and placed at 37 8C for 12–24 h.
explant and BAP concentration. The experiment was carried out with 15replicates and a total of 300 explants.
Molecular analysis was performed on putative transgenic
conditions were established first to efficiently regenerate R.
and control plants by PCR and Southern hybridisation.
graveolens plants. Secondly, transformation conditions were
Qiagen ‘DNeasy plant maxi kit’ was used to isolate genomic
determined using an Agrobacterium binary vector method
DNA. For PCR analysis, a 552 bp gus sequence was
with the objective to get the highest percentage of
amplified using the following primer sets: 50-ATG GTC
transformed plant. Finally, histochemical and molecular
CGT CCT GTA GAA ACC CCA ACC CGT GAA ATC-30
techniques were used to verify that plants were genetically
and 50-CGC AGC GTA ATG CTC TAC ACC ACG CCG
AAC ACC TGG GTG GAC GAT A-30. The PCR conditionswere: 95 8C for 5 min followed by 35 cycles at 95 8C for
60 s, 65 8C for 30 s, and 72 8C for 40 s. Amplified DNA wasanalysed on a 1% (w/v) TAE agarose gel.
Regeneration ability of R. graveolens L. was tested by
Southern blot was performed on eight randomly chosen
cultivating five types of explants (apices, cotyledons,
KANR plants and one wild (control) plant. Each Southern blot
hypocotyls, roots and root tips) on MS(30) containing
analysis was carried out on 1 g of DNA extracted from the
different concentrations of BAP (0, 0.1, 1 and 2 mg/l). After
leaves using Qiagen ‘DNeasy plant maxi kit’. DNA (20 mg)
2 weeks, hypocotyls gave the highest number of regenerated
from each sample were individually digested for 20 h at 37 8C
shoots (). A maximum of three to four shoots per
with a mix of three enzymes (150 units of EcoRI, 150 units of
hypocotyls were regenerated on MS(30) containing 0.1 mg/l
BamHI and 70 units of BglII) added in two times. There is no
BAP When reaching a height of 1 cm, these shoots
restriction site for these three enzymes in the gus sequence, so
were transferred in glass vessels containing MS(30) with
the number of insertions of the gus gene in the genome of R.
IAA (0, 0.1, 0.5 and 1 mg/l) for rooting. Shoots rooted better
graveolens L. is equal to the number of fragments obtained.
when they were grown on MS(30) without IAA than after
The digested DNA was precipitated with 0.8 vol isopropanol
addition of this compound. After 6 weeks, 55% of the shoots
in presence of 0.3 M AcNa pH 5.6. Pellets were resuspended
had four roots a least, enough to transfer them into a
in TAE buffer. Prior to loading on a 0.8% agarose gel, we
added loading buffer (TAE, 1Â; 50% glycerol; 0.1%
Hypocotyls appeared to regenerate plants most effi-
Bromophenol Blue; 0.1 SDS) and heated the samples for
ciently. These explants were retained for further genetic
10 min at 65 8C. Digested DNA was blotted on a positively
Bioscience). The hybridisation reaction was carried out for16 h at 65 8C using as a probe the gus PCR product obtainedwith the primer set described above and the pBin-gus plasmid.
Table 1Effect of IAA concentration on shoots rooting
This probe was labelled with 32P using Ready-To-GoTMLabelling Beads (-dCTP) according to the manufacturer’s
recommendation (Amersham-Bioscience).
Shoots (1 cm-high) were grown on agar-solidified MS(30) medium contain-ing IAA (0, 0.1, 0.5 and 1 mg/l). After 6 weeks, the percentage of shoots
Regeneration of a plant is a prerequisite to further
having at least four roots was evaluated. The experiments were carried out
transformation. Therefore, plant tissue and organ culture
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
Table 2Effects of acetosyringone concentration (0, 50 and 250 mM) in the co-culture medium of Ruta graveolens L. and Agrobacterium tumefaciens
Selection method was done in one step (with 50 mg/l kanamycin) or two step (with 75 mg/l kanamycin). ND: not determined.
a Number of Ruta graveolens L. hypocotyls infected with Agrobacterium tumefaciens. b SE: Percentage of KANR plants with the gus gene (PCR detection). c Percentage of KANR plants with GUS activity detected by histochemical assays. d TE: Transformation efficiency, number of explants that regenerated at least one transformed plant confirmed by PCR on gus gene.
3.2. Genetic transformation and regeneration of plants
protocol by comparing TE obtained with the ‘one step’ and‘two step’ selection method. However, we had previously
established that numerous plants cannot support a strong and
Phenolics like acetosyringone are well-known virulence
early selection, therefore, KAN concentration in the ‘one
inducers for Agrobacterium The aim of the first
step’ selection method was diminished from 75 to 50 mg/l.
transformation experiment was to determine the optimum
The percentage of KANR plants obtained with the two step
concentration of acetosyringone in co-culture medium.
selection method (14.2%, ) was 1.5 folds the one
Agrobacterium tumefaciens and hypocotyls were co-
recorded with the one step selection method (9.2%,
cultivated for 3 days on MS(30)BAP0.1 supplemented with
Consequently, the one step selection method was not used
0, 50 or 250 mM acetosyringone. Subsequently, the
hypocotyls were treated as previously described for the‘two step’ selection method. Explants consisting of 1 cm of
3.2.4. Optimised protocol (two step selection method)
developing shoot explants were then transferred to a fresh
Transformation efficiency is above 11.1% for routine
selection medium containing KAN 75 mg/l for 3 weeks and
experiments and could reach 22.2% in some experiments.
could reach 2–3 cm at the end of this period. When
Because KANR plants obtained in the different transforma-
transferred to a fresh MS(30) medium without antibiotic for
tion experiments are coming from different hypocotyls, all
6–8 weeks, they could grow and root. Results shown in
transgenic plants are originating from different transforma-
demonstrate that the transformation of R. graveolens
tion events. After selection screening, the shoots that showed
L. was improved in the presence of acetosyringone, since
good root development were grown ex vitro into a
14.2% of the initial explants produced KANR plants when
phytotronic chamber. All of them exhibited normal
250 mM of acetosyringone was added to the co-culture
development as compared to control plants.
medium. Selection efficiency (SE) was established at 78%for this acetosyringone treatment.
3.3. Molecular analysis of transgenic plants
PCR analysis was carried out to confirm the presence of
Pre-culture of 0, 1 or 3 days was tested in order to
the gus gene in the genome of putative transformants
improve the transformation efficiency. Hypocotyls were
(This was performed on 21 out of the 27 KANR
pre-cultured on MS(30)BAP0.1 before being co-cultured
plants. The 550 bp expected gus fragment was found in the
with A. tumefaciens on MS(30)BAP0.1 with 250 mM
positive control (pBin-gus plasmid) as well as in 78% of the
acetosyringone. Selection with 75 mg/l KAN was used
analysed plants. Wild plants (controls) did not show the
as previously described. This experiment was duplicated.
The transformation efficiency (TE) was about 5%, whateverthe time of pre-culture. Consequently, even if thetransformation rate of these experiments is lower thanthose described previously, it demonstrates that pre-culturehas no positive effect for genetic transformation of R. graveolens L.
Fig. 2. PCR analysis of putative transformed plants. Genomic DNAs wereamplified with primers corresponding to a 550 bp gus gene sequence. Lanes:
Decreasing the number of steps necessary to obtain
PM, molecular weight ladder; (+) positive control (pBin-gus); (À) negative
transgenic plants would speed up genetic transformation of
control (wild type plant); lanes 1, 2, 4–8, transformed plants; lane 3,
R. graveolens L. Therefore, we tested a more simple
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
300 mg/l cefotaxime, or in two steps on MS(30)BAP0.1supplemented with 300 mg/l cefotaxime and 75 mg/lkanamycin. Stable transgene integration was confirmedby growth on selection medium for nptII, by PCR andSouthern blot analyses, and histochemical revelation of GUSactivity. Results observed with acetosyringone suggest thatthis compound is useful to improve genetic transformationof R. graveolens L. as for many other species It hasbeen demonstrated that a two step selection protocolincreases the transformation efficiency and minimizes theincidence of escapes even with a higher concentration ofselective agent The remaining escapes could beexplained either by Agrobacterium endogenous contamina-tion of plant tissues, or by a low transformation frequencyversus a high efficiency of shoot formation. In this lattercase, the non-transformed cells could be protected by thesurrounding transformed cells resulting in chimeric shoots. Pre-culture of hypocotyls, prior to bacterial co-incubation,
Fig. 3. Southern blot analysis of putative transformed plants. Genomic
had no positive effect on transformation efficiency.
DNAs were digested with a mix of three enzymes (EcoRI, BamHI and BglII)
Starting from the original seeds, it is possible to obtain
and probed using a 32P labelled GUS probe, prepared from the plasmid
transgenic plants growing on soil within 4–5 months.
pBin-gus. Lane wt: wild type control plant, lanes 1–8: transformed plants. Dots indicate insertion events.
Because the original plant material is composed of seeds, itprevents possible somaclonal variations associated with longlasting in vitro culture of callus.
Southern blot analysis was done in order to estimate the
R. graveolens L. produces many different secondary
number of inserted loci. DNA from eight plants were
metabolites. The efficiency of this transformation protocol
analysed. shows the Southern blot hybridisation
will allow us to easily generate mutants up or down
pattern obtained for the gus probe. Three out of eight
regulated for various key-enzymes involved in secondary
analysed plants showed a single insert of the gus gene (lines
pathways. This will help us to understand secondary
3, 4 and 7), while four lines (2, 5, 6, 8) showed two
metabolites regulations, especially in the furocoumarin
hybridisation bands for the gus gene, and line 1 has three
pathway. A new set of mutants has been recently produced
copies. No band was observed in the control plant.
with a modified cinnamate-4-hydroxylase (C4H) expressionunder the control of the CaMV 35S promoter using this
About 67% of the plants transformed with the gus gene
expressed b-glucuronidase (). Staining for GUS
expression revealed variable colour intensities in trans-formed plants. No GUS staining was observed in control
We acknowledge the Re´gion Lorraine for financial
support of the research program (PRST Bioinge´nierie) andthe French Ministry of Research for Ph.D. grant (KarineLie`vre). We thank Paul Lacoste and Se´bastien Doerper for
An efficient Agrobacterium-mediated transformation
protocol for R. graveolens L. is presented for the first time.
The only related studies are those of Porter and Eilert who infected R. graveolens with wild type Agrobacterium
[1] E. San Miguel, Rue (Ruta L., Rutaceae) in traditional Spain: frequency
rhizogenes or tumefaciens. However, these authors neither
and distribution of its medicinal and symbolic applications, Econ. Bot.
tried to genetically transform R. graveolens L. with
engineered Agrobacterium strains, nor to genetically/
[2] A.M. Zobel, S.A. Brown, Histochemical localization of furanocou-
biochemically confirm the status of the transformants.
marins in Ruta graveolens shoots, Can. J. Bot. 67 (1989) 915–921.
Our transformation procedure is based on a 3 days co-culture
[3] P.S. Song, K.J. Tapley, Photochemistry and photobiology of psoralens,
Photochem. Photobiol. 29 (1979) 1177–1197.
of seedling hypocotyls with A. tumefaciens C58C1RifR in
[4] A. Gravot, R. Larbat, A. Hehn, K. Lievre, E. Gontier, J.L. Goergen, F.
the presence of 250 mM acetosyringone. Regeneration and
Bourgaud, Cinnamic acid 4-hydroxylase mechanism-based inactiva-
selection were obtained in one step on MS(30)BAP0.1 with
tion by psoralen derivatives: cloning and characterization of a C4H
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
from a psoralen producing plant-Ruta graveolens-exhibiting low
for Agrobacterium-mediated gene transfer to plants, Nucleic Acids
sensitivity to psoralen inactivation, Arch. Biochem. Biophys. 422
[9] P. Dion, C. Belanger, D. Xu, M. Mohammadi, Effect of acetosyringone
[5] U. Eilert, Elicitor induction of secondary metabolism in dedifferen-
on growth and oncogenic potential of Agrobacterium tumefaciens,
tiated in vitro systems of Ruta graveolens, in: W.G.W. Kurz (Ed.),
Methods Mol. Biol. 44 (1995) 37–45.
Primary and Secondary Metabolism of Plant, 1989.
[10] S. Suzuki, M. Nakano, Agrobacterium-mediated production of trans-
[6] J.R. Porter, Host range and implications of plant infection by
genic plants of Muscari armeniacum Leichtl. ex Bak, Plant Cell Rep.
Agrobacterium rhizogenes, Crit. Rev. Plant Sci. 10 (4) (1991) 387–
[11] E. Sales, J. Segura, I. Arrillaga, Agrobacterium tumefaciens-mediated
[7] T. Murashige, F. Skoog, A revised medium for rapid growth and
genetic transformation of the cardenolide-producing plant Digitalis
bio assay with tobacco tissues cultures, Physiol. Plant. 15 (1962)
minor L., Planta Med. 69 (2) (2003) 143–147.
[12] J.L. Yao, D. Cohen, R. Atkinson, K. Richardson, B. Morris, Regen-
[8] R. Deblaere, B. Bytebier, H. De Greve, F. Deboeck, J. Schell, M. Van
eration of transgenic plants from the commercial apple cultivar Royal
Montagu, J. Leemans, Efficient octopine Ti plasmid-derived vectors
Gala, Plant Cell Rep. 14 (1995) 407–412.
AIMAC 2005 8th International Conference on Arts and Cultural Management July 3-6, 2005. Montréal, Canada Sunday, July 3rd 18:00 Welcome Cocktail: Musée du Château Ramezay 280 Notre-Dame Street East Monday, July 4th Opening remarks "We Manage as We Are" by Laurent Lapierre Session 1 - Consumer Behaviour Room: Meloche Monnex Chair : Anne-Gaëlle Jol
International Journal of Medical Informatics 51 (1998) 59 – 68MERIT-9: a patient information exchange guideline usingMichio Kimura a,*, Kazuhiko Ohe b, Hiroyuki Yoshihara c, Yutaka Ando d,Fumiaki Kawamata e, Fumito Tsuchiya f, Hiroyuki Furukawa g, Shingo Horiguchi b,Takaya Sakusabe h, Shigeki Tani a, Masanori Akiyama ia Department of Medical Informatics , School of Medicine , Hamamat


 Genetic transformation of the medicinal plant Ruta graveolens L.
Genetic transformation of the medicinal plant Ruta graveolens L.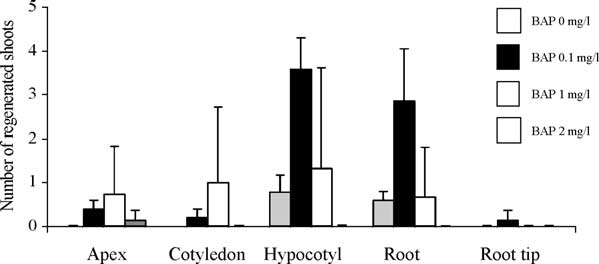 K. Lie`vre et al. / Plant Science 168 (2005) 883–888
to ensure that all the transformed shoots were coming fromindependent transformation events.
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
to ensure that all the transformed shoots were coming fromindependent transformation events.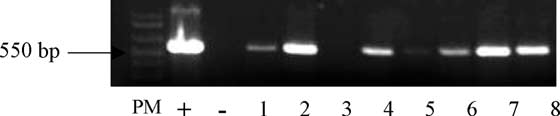 K. Lie`vre et al. / Plant Science 168 (2005) 883–888
Table 2Effects of acetosyringone concentration (0, 50 and 250 mM) in the co-culture medium of Ruta graveolens L. and Agrobacterium tumefaciens
Selection method was done in one step (with 50 mg/l kanamycin) or two step (with 75 mg/l kanamycin). ND: not determined.
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
Table 2Effects of acetosyringone concentration (0, 50 and 250 mM) in the co-culture medium of Ruta graveolens L. and Agrobacterium tumefaciens
Selection method was done in one step (with 50 mg/l kanamycin) or two step (with 75 mg/l kanamycin). ND: not determined.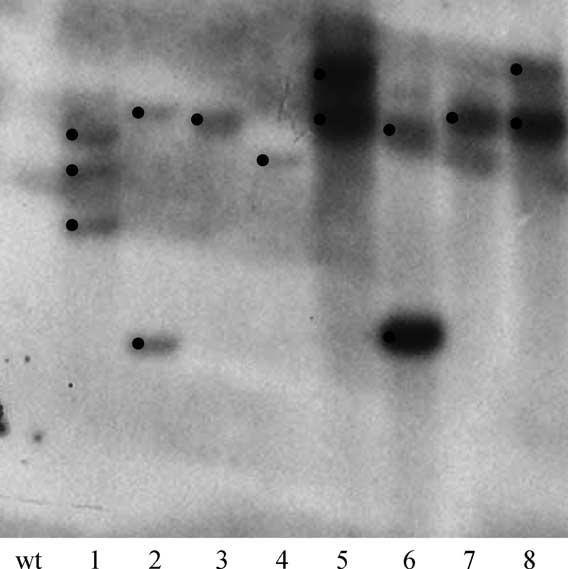 K. Lie`vre et al. / Plant Science 168 (2005) 883–888
300 mg/l cefotaxime, or in two steps on MS(30)BAP0.1supplemented with 300 mg/l cefotaxime and 75 mg/lkanamycin. Stable transgene integration was confirmedby growth on selection medium for nptII, by PCR andSouthern blot analyses, and histochemical revelation of GUSactivity. Results observed with acetosyringone suggest thatthis compound is useful to improve genetic transformationof R. graveolens L. as for many other species It hasbeen demonstrated that a two step selection protocolincreases the transformation efficiency and minimizes theincidence of escapes even with a higher concentration ofselective agent The remaining escapes could beexplained either by Agrobacterium endogenous contamina-tion of plant tissues, or by a low transformation frequencyversus a high efficiency of shoot formation. In this lattercase, the non-transformed cells could be protected by thesurrounding transformed cells resulting in chimeric shoots.
K. Lie`vre et al. / Plant Science 168 (2005) 883–888
300 mg/l cefotaxime, or in two steps on MS(30)BAP0.1supplemented with 300 mg/l cefotaxime and 75 mg/lkanamycin. Stable transgene integration was confirmedby growth on selection medium for nptII, by PCR andSouthern blot analyses, and histochemical revelation of GUSactivity. Results observed with acetosyringone suggest thatthis compound is useful to improve genetic transformationof R. graveolens L. as for many other species It hasbeen demonstrated that a two step selection protocolincreases the transformation efficiency and minimizes theincidence of escapes even with a higher concentration ofselective agent The remaining escapes could beexplained either by Agrobacterium endogenous contamina-tion of plant tissues, or by a low transformation frequencyversus a high efficiency of shoot formation. In this lattercase, the non-transformed cells could be protected by thesurrounding transformed cells resulting in chimeric shoots.